Диносаурус
Диносаурус — група влекачи[заб. 1] од кладот „Диносаурија“ (Dinosauria). Тие првпат се појавиле за време на геолошкиот период тријас, помеѓу 243 и 233,23 милиони години, иако точното потекло и времето на еволуцијата на диносаурусите е предмет на постојани истражувања. Тие станале доминантни копнени ’рбетници пред 201,3 милиони години и нивната доминација продолжила во текот на периодот Јура и Креда. Фосилните записи покажуваат дека птиците се современи пердувести диносауруси, кои еволуирале од претходните тероподи за време на доцната јура. Како такви, птиците биле единствената лоза на диносауруси што го преживеале исчезнувањето во кредно-палеогенскиот настан на изумирање пред околу 66 милиони години. Затоа, диносаурусите можат да се поделат на птичји диносауруси и нептичји диносауруси.
| Диносауруси Период: доцен тријас - доцна креда денес се останати нивните потомци, птиците | |
|---|---|

| |
| Скелети на тираносаурус (лево) и апатосаурус (десно) | |
| Научна класификација [ у ] | |
| Царство: | Животни |
| Тип: | Хордови |
| Класа: | Влекачи |
| (нерангирано): | Орнитодири |
| (нерангирано): | Диносаурусолики |
| (нерангирано): | Диносаурусовидни |
| Надред: | Диносауруси Овен, 1842 |
| Редови и подредови | |
| |
Диносаурусите се разновидна група на животни од таксономска, морфолошка и еколошка гледна точка. Птиците, со над 10.000 живи видови, се најразновидната група на ’рбетници покрај перциформните риби. Користејќи докази за фосили, палеонтолозите идентификувале над 500 различни родови и повеќе од 1.000 различни видови нептичји диносауруси. Диносаурусите живееле на секој континент а тоа се докажува од принајдените фосилни остатоци и преку постоечките видови птици. Во текот на првата половина на XX век, пред птиците да бидат препознаени како диносауруси, поголемиот дел од научната заедница верувала дека диносаурусите биле слаби и ладнокрвни. Повеќето истражувања спроведени од 1970-тите укажале на тоа дека сите диносауруси биле активни животни со зголемен метаболизам и бројни прилагодувања за социјална интеракција. Некои биле тревојадни, други месојади. Доказите сугерираат дека сите диносауруси гнезделе јајца, и таа градба на гнездо била одлика што ја делеле многу диносауруси, и птичји и нептичји.
Додека диносаурусите биле двоножни, многу изумрени групи вклучувале четириножчни видови, а некои имале можност и да комбинираат. Елаборатните структури на предниот дел, како рогови на пример, се заеднички за сите групи на диносауруси, а некои изумрени групи развиле скелетни модификации како на пример коскени оклопи. Додека денешната преживеана птичја лоза на диносаурусите е генерално мала поради ограничувањата на летот, многу праисториски диносауруси (нептичји и птичји) биле со големо тело - најголемите диносауруси од групата сауроподи (или гуштероноголики) се проценува дека достигнувале 39,7 метри и височина од 18 метри и истите претставувале најголеми копнени животни на сите времиња. Сепак, идејата дека нептичјите диносауруси биле подеднакво гигантски е заблуда заснована делумно на пристрасноста кон зачувување, бидејќи видовите со големи и цврсти коски имаат поголема веројатност да траат сè додека не бидат фосилизирани. Многу диносауруси биле прилично мали: Xixianykus, на пример, бил долг само околу 50 сантиметри.
Од наоѓањето и признавањето на првите фосили на диносауруси на почетокот на 19 век, монтираните фосилни скелети на диносауруси биле главна атракција во музеите ширум светот, а диносаурусите станале траен дел од светската култура. Големината на некои групи диносауруси, како и нивната навидум монструозна и фантастична природа, обезбедиле редовно појавување на диносаурусите во најпродавани книги и филмови, како што е „Паркот Јура“. Постојаниот ентузијазам во јавноста за животните резултирал со значително финансирање на науката за диносаурусите, а новите откритија редовно ги покриваат медиумите.
Етимологија
уреди
Таксонот „Dinosauria“ (Диносаурија) бил официјално именуван во 1841 година од страна на палеонтологот Сер Ричард Овен, кој термин го користел за разликување од сауриските влекачи, кои тогаш биле признати во Англија и низ целиот свет.[1][2] Терминот е изведен од старогрчкиот збор δεινός (deinos), што значи „страшен, моќен или страшно голем“ и σα andρος (sauros), што значи „гуштер или влекач“[1][3]. Иако таксономското име често се толкува како упатување на забите, канџите и другите застрашувачки одлики на диносаурусите, Овен имал за цел само да ги оцрта нивната големина и величественост[4].
Другите праисториски животни, вклучително птеросаури, мозасаури, ихтиосаури, плезиосаури и Диметредон, иако честопати популарно ги замислувале како диносауруси, не се класифицирани таксономски како диносауруси[5]. Птеросаурусите се далечно поврзани со диносаурусите, кои се членови на Орнитодира. Другите споменати групи се, како диносаурусите и птеросаурусите, членови на Сауропсида, освен Диметредон (кој е синапсид).
Поим
уредиОд филогенетската номенклатура, диносаурусите обично се дефинираат како група која се состои од најблискиот древен заеднички предок (MRCA) на Трицератопс и современите птици (Neornithes) и сите нивни потомци[6]. Исто така, било предложено Диносаурија да се дефинира во однос на МРЦА на Мегалосаурус и Игуандон, бидејќи тие биле два од трите рода цитирани од Ричард Овен кога била признена Диносаурија[7]. И двете дефиниции резултираат во ист сет на животни кои се дефинираат како диносауруси: „Dinosauria = Ornithischia + Saurischia“, опфаќајќи анкилосаури (оклопни тревојади четириножни), стегосаури (тревојадни четириножни), цератопсиди (тревојадни со рогови) или четириножни тревојади), Тероподи (претежно двоножни месојади и птици) и сауроподоморфи (претежно големи тревојадни четириножни со долги вратови и опашки)[8].
Птиците денес се признаваат како единствена преживеана лоза на тероподни диносауруси. Во традиционалната таксономија, птиците се сметаат за посебна класа кои еволуирале од диносаурусите. Сепак, мнозинството современи палеонтолози кои се занимаваат со диносаурусите го отфрлаат традиционалниот стил на класификација во корист на филогенетската таксономија; овој пристап бара, за да биде група природна, сите потомци на членовите на групата да мора да бидат вклучени во групата. Така, птиците се сметаат за диносауруси, а диносаурусите не се истребани[9]. Птиците се класифицирани како припадници на подгрупата Манираптора, кои се целуросаури[10].
Истражувањето на Метју Г. Барон, Дејвид Б. Норман и Пол М. Барет во 2017 година покажало радикална ревизија на диносауриската систематика. Филогенетска анализа од Барон ја обновил Орнитишија (Птицоколковни) како поблиска до терододите од сауроподоморфите, наспроти традиционалното соединување на тероподите со сауроподоморфите. Самата Диносаурија била редефинирана како последен заеднички предок на Triceratops horridus, Passer domesticus и Diplodocus carnegii и сите нејзини потомци, за да се осигура дека сауроподите и роднините остануваат вклучени како диносауруси[11][12].
Генерален опис
уредиКористејќи една од горенаведените дефиниции, диносаурусите генерално може да се опишат како архосауруси со задни екстремитети држени исправени под телото[13]. Многу праисториски животински групи популарно се замислуваат како диносауруси, како што се ихтиосаури, мозаури, плезиосаури, птеросаури и пеликосаури (особено Диметродон), но не се класифицирани научно како диносауруси и ниту една група од овие животни немале држечки став карактеристичен за вистинските диносауруси[14]. Диносаурусите биле доминантни копнени ’рбетници во мезозојската ера, особено во периодот Јура и Креда. Другите групи животни биле ограничени по големина. Цицачите, на пример, ретко ја надминувале големината на домашна мачка и генерално биле месојади со големина на глодачи[15].
Диносаурусите отсекогаш биле исклучително разновидна група животни. Според една студија од 2006 година, досега се идентификувани над 500 родови нептичји диносауруси, а вкупниот број на зачувани родови во фосилните записи е проценет на околу 1850, според кој податок останува да се откријат уште скоро 75% од видовите[16]. Претходната студија предвидувала дека постоеле околу 3.400 родови на диносауруси, вклучително и многу кои не би биле зачувани во фосилните записи[17]. До 17 септември 2008 година биле именувани 1.047 различни видови диносауруси
.
Во 2016 година, проценетиот број на видови диносауруси што постоеле во мезозоикот се проценува на 1.543-2.468[18][19]. Некои се тревојадни, други месојади, вклучително и јадечи на семе, риба, инсективатори и сештојади. Додека предците на диносаурусите биле двоножни (како и сите современи птици), некои праисториски видови биле четириножни, а други, како Анхисаурус и Игуанодон, можеле да одат лесно на две или четири нозе. Мзозчните модификации како рогови и гребени се вообичаени одлики на диносаурусите, а некои изумрени видови имале коскени оклопи. Иако се познати по големи димензии, многу мезозоични диносауруси биле со човечка големина или помали, а современите птици се генерално мали по големина. Диносаурусите денес живеат на секој континент, а фосилите покажуваат дека тие постигнале глобална распространетост барем во ерата на раниот период на Јура[20]. Современите птици живеат во повеќето достапни живеалишта, од копнени до морски и има докази дека некои не-птичји диносауруси може да летаат или барем да лазат, а други, како што се спиносауриди, имале полуакватични навики[21].
Разликување на анатомски одлики
уредиДодека неодамнешните откритија го отежнувале презентирањето на универзално договорениот список на карактеристични одлики на диносаурусите, скоро сите досега откриени диносауруси споделуваат одредени модификации на скелетот на предците од архосаурискиот скелет, или се јасни потомци на постарите диносауруси кои ги покажуваат овие модификации. Иако некои подоцнежни групи диносауруси имале дополнително изменети верзии на овие црти, тие се сметаат за типични за Диносаурија, кои ги имале најраните диносауруси и им ги пренеле на нивните потомци. Таквите модификации, кои потекнуваат од најновиот заеднички предок на одредена таксономска група, се нарекуваат синапоморфии на таква група[22].
Деталната проценка на меѓусебните врски на архосаурусот од страна на Стерлинг Несбит[23] потврдила дванаесет недвосмислени синапоморфии. Тој пронашол и голем број на понатамошни потенцијални синапоморфии. Некои од нив се исто така присутни во силесауридите, кои Несбит ги опоравил како сестринска група на Диносаурија.

Разновидни други одлики на скелетот ги споделуваат диносаурусите. Меѓутоа, бидејќи тие се или заеднички за другите групи на архосаури, или не биле присутни кај сите рани диносауруси, овие одлики не се сметаат за синапоморфии. На пример, како диапсиди, диносаурусите имале претходно два пара инфрастемпорални фенестри (отвори во черепот зад очите), и како членови на групата Архосаурија, имале дополнителни отвори во устата и долната вилица[24]. Дополнително, се знае дека неколку одлики за кои некогаш се сметало дека се синапоморфии, се појавиле пред диносаурусите или биле отсутни кај најраните диносауруси и независно се развиле од различни групи на диносауруси. Овие вклучуваат издолжена плешка, крсна коска составена од три или повеќе споени пршлени (три се наоѓаат кај некои други архосаури, но само два се наоѓаат кај Херерасаурус)[6]; и acetabulum (карлича чаша), или колк, со дупка во центарот на нејзината внатрешна површина[25][26]. Друга тешкотија при утврдување на изразито диносауриски одлики е тоа што раните диносауруси и другите архосауруси од ерата на доцниот Тријас честопати се слабо познати и биле слични на многу начини. Овие животни понекогаш биле погрешно идентификувани во литературата[27]

Диносаурусите стојат со задните екстремитети исправени на начин сличен на повеќето современи цицачи, но различен од повеќето други влекачи, чии екстремитети се шират на двете страни[28]. Ова држење на телото се должи на развојот на странично свртената вдлабнатина во карлицата и соодветна внатрешно свртена изразена глава на бедрената коска[29]. Нивната исправена положба им овозможувала на раните диносауруси да дишат лесно додека се движат[30].
Еволутивна историја
уредиПотекло и рана еволуција
уредиДиносаурусите се разминале од нивните предци архосаурите за време на доцниот Тријас, околу 20 милиони години во Пермско-тријаското изумирање кога околу 96% од сите морски видови и 70% од копнените ’рбетници пред околу 252 милиони години исчезнале[31][32]. Радиометричкото датирање на карпестата формација што содржела фосили од раниот род на диносауруси Еораптор на возраст од 231,4 милиони години, го утврдува неговото присуство во фосилните пронајдоци од тоа време[33] Палеонтолозите сметаат дека Еорапторот наликува на заеднички предок на сите диносауруси[34]. Доколку ова е вистина, неговите одлики укажуваат на тоа дека првите диносауруси биле мали, двоножни грабливци[35]. Откривањето на примитивни, слични на диносауруси видови како што се Марасухус и Лагерпетон во аргентинските среднотријаски слоеви го поддржува ова гледиште. Анализата на пронајдени фосили сугерира дека овие животни биле навистина мали и биле двоножни грабливци. Диносаурусите може да се појавиле уште пред 243 милиони години, за што сведочат остатоци од родот Њасасаурус (Nyasasaurus) од тој период, иако познатите фосили на овие животни се премногу фрагментарни за да се каже дали се диносауруси или многу блиски роднини на диносаурусите[36]. Палеонтологот Макс Ц. Лангер (2018) утврдил дека Стаурикосаурусот од формацијата Санта Марија датира од пред 233,23 милиони години, што го прави постар во геолошка возраст од Еораптор[37].
Кога се појавиле диносаурусите, тие не биле доминантни копнени животни. Терестријалните живеалишта биле окупирани од разни видови архосауроморфи и терапсиди, како цинодонти и ринхосаури. Нивните главни конкуренти биле псевдосаурите, како што се етосаурусите, орнитосаурусите, кои биле поуспешни од диносаурусите[38]. Повеќето од овие други животни изумреле во тријасот, во еден од двата настани на истребување. Прво, пред околу 215 милиони години, исчезнале различни базални архосауроморфи, вклучувајќи ги и проторосаурусите. Ова било проследено со настанот на истребување во Јура (пред околу 201 милиони години), каде исчезнале повеќето други групи на рани архосаури, како етосаури, фитосауруси итн. Ринхосаурусите и дицинодонтите преживеале (во некои области) барем во почетокот на средниот норијски и доцниот норијски или најраниот ретски период, соодветно[39][40], и точниот датум на нивното истребување е неизвесен. Овие загуби оставиле зад себе копнена фауна на крокодиломорфи, диносауруси, цицачи, птеросаури и желки[6]. Првите неколку линии на рани диносауруси се диверзифицирале низ фазите карник и норик на тријасот[8]. Исто така, забележително, имало и зголемена стапка на истребување за време на Карвијанскиот настан[41].
Еволуција и палеобиогеографија
уредиЕволуцијата на диносаурусите по периодот Тријас ги следи промените во растителноста и местоположбата на континентите. Во доцниот Тријас и раната Јура, континентите биле поврзани меѓусебно, образувајќи го суперконтинентот Пангеја и постоела светска фауна на диносауруси, претежно составена од месојади целофизоиди и рани тревојади составени од сауроподоморфи[42]. Голосемените растенија претставувале потенцијален извор на храна кон крајот на тријасот. Раните сауроподоморфи немале софистицирани механизми за обработка на храната во устата, и затоа морало да користат други средства за распаѓање на храната долж дигестивниот тракт[43]. Општата хомогеност на диносауриските фауни продолжила во средната и доцната Јура, каде што повеќето имале грабливци кои се состоеле од цератосаури, спиносауроиди и карносаури, и тревојади што се состоеле од Ornithischia и големи сауроподи. Примери за ова ја вклучуваат Морисоновата формација на Северна Америка и формацијата Тендагуру на Танзанија. Диносаурусите во Кина покажуваат некои разлики, со специјализирани синопрапторидни тероподи и невообичаени сауроподи со долг врат како Маменчисаурус[42]. Анкилосаурусите и орнитоподите исто така станале сè почести, но платеосауридите изумреле. Четинарите и птеридофитите биле најчестите растенија. Сауроподите, како и претходните просауроподи, немале орални процеси, но орнитоподите развивиле разни средства за справување со храна во устата, вклучувајќи потенцијални органи слични на образите за да ја задржат храната во устата и движења на вилицата за џвакање на храната[43]. Друг значаен еволутивен настан на Јура била појавата на вистински птици, кои потекнуваат од манирапторските целуросауруси[10].

Со периодот на раната креда и тековното распаѓање на Пангеја, диносаурусите станале силно диференцирани од копното. Во најраниот дел од ова време, се ширеле анкилосаурусите, игуандонтите и брахиосаурусите низ Европа, Северна Америка и Северна Африка. Подоцна, овие биле надополнети или заменети во Африка со големи спиносауроиди и кархародонтосаурите тероподи и ребахисауриди и титаносауриски сауроподи, пронајдени во Јужна Америка. Во Азија, манирапторанските целуросауруси, како што се дромеосауридите, тродонтидите и овирапторосаурите, станале вообичаени тероподи, а анкилосаурусите и раните цератопсиди како Пситакосаурус станале важни тревопасни животни. Во меѓувреме, Австралија била дом на фауна на базални анкилосаури, хипсилофодонти и игуандонти[42]. Изгледа дека стегосаурусите исчезнале во одреден момент во доцната рана креда или раната доцна креда. Голема промена која се слуќила во раниот период на креда и се засилила во доцната, е еволуцијата на цветните растенија. Во исто време, неколку групи тревојади на диносауруси еволуирале и наметнале пософистицирани начини за орална обработка на храната. Цератопсидите развиле метод на мелење со забите наредени едни на други[43]. Некои сауроподи, исто така, еволуирале со своите заби, а најдобар пример за тоа е Нигеросаурусот[44].

Во доцната креда имало три фауни на општите диносауруси. На северните континенти на Северна Америка и Азија, главните тероподи биле тираносауридите и разни видови помали манирапторски троподи, со доминантно орнитхиски тревојади, хадросауриди, цератопси, анкилосауриди и пахицефалосауриди. На јужните континенти што ја сочинувале веќе разделената Гондвана, абелисауридите биле обични тероподи, а титаносауриските сауроподи - обичните тревопасни животни. Во Европа преовладувале дромаеосауридите, рабдодонтидите, игуаодонтите, нодосауридните анкилосауруси и титаносауриските сауроподи[42][45] . Цветните растенија во голема мера биле распространети, со првите треви кои се појавиле до крајот на периодот креда. Поради начинот на исхрана, хадросауридите станале исклучително разновидни низ Северна Америка и Азија. Троподите делувале и како тревојади или сештојади, со тоа што станале вообичаени теризиносауруси и орнитомимосаури[43].
Настанот на исчезнување креда-палеоген, што се случил пред околу 66 милиони години на крајот на од периодот креда, предизвикал истребување на сите групи диносауруси, освен на птиците неорнитин. Некои други групи на дијапсиди, како што се крокодили, желки, гуштери, змии и хористиодерани, исто така, го преживеале настанот[46].
Преживеаните лози на неортински птици, вклучувајќи ги и предците на современите ратити, патки и кокошки и разни водни птици, диверзифицирале брзо на почетокот на палеогенскиот период, влегувајќи во еколошките ниши оставени празни со истребување на мезозојските диносаурурски групи како што се енанциорнисовите птици, водни хесперорнитини, па дури и поголеми копнени тероподи. Често се наведува дека цицачите се натпреварувале надвор од неортините за доминација на повеќето копнени ниши, но многу од овие групи коегзистирале со богати фауни на цицачи во поголемиот дел од ерата на кенозоикот[47]. Фуроратосовите птици биле месојадни заедно со грабливите цицачи[48][49], и ратитите сè уште биле прилично успешни како тревојади од средна големина[50].
Класификација
уредиДиносаурусите припаѓаат на група позната како архосауруси, во која спаѓаат и современите крокодили. Во рамките на групата архосауруси, диносаурусите се разликуваат најзабележително според одењето. Нозете на диносаурусите се протегаат директно под телото, додека нозете на гуштерите и крокодилите се протегаат на двете страни[22].
Колективно, диносаурусите се поделени во две примарни гранки, Сауришија и Орнитишија. Првите ги вклучуваат таксоните кои имаат понов заеднички предок со птиците отколку со Орнитишија, додека Орнитишија ги вклучува сите таксони кои имаат понов заеднички предок со Трицератопс отколку со Сауришија. Анатомски, овие две групи можат да се разликуваат најзабележително според нивната карлична структура. Раните сауроподи ја задржале структурата на колкот на нивните предци, со препонска коска насочена кранијално или напред[29][51]. Оваа основна форма била изменета со вртење на препонската коска наназад во различни степени во неколку групи (Херерасаурус[52], теризиносаурус)[53]. Сауришија вклучува тероподи (исклучиво двоножни) и сауроподоморфи (тревојади со долг врат, кои вклучуваат четириножни групи)[21][54].
Спротивно на тоа, Орнитишија - „птици“, имале карлица што површно личи на карлицата на птицата: препонската коска била ориентирана кон опашката (насока наназад). За разлика од птиците, оваа коска обично имашла и дополнителен процес на насочување нанапред. Орнитишија вклучува различни видови кои првенствено биле тревојади. И покрај поимите „колк на птица“ и „колк на гуштер“, птиците не се дел од Орнитишија, туку од Сауришија - птиците еволуирале од претходните диносауруси со „колкови на гуштер“[22].
-
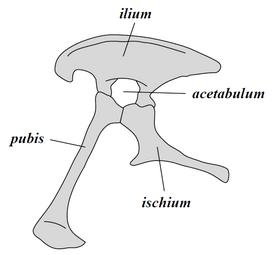 Карлица на Сауришија
Карлица на Сауришија -

-
 Карлица на Орнитишија
Карлица на Орнитишија -





Таксономија
уредиПодолу е поедноставена класификација на групи на диносауруси врз основа на нивните еволутивни врски и организирани врз основа на списокот на видови мезозоични диносауруси обезбедена од Холц (2007). Подетална верзија може да се најде на Класификација на диносаурусите. Ознаката (†) се користи за означување групи без живи членови.
- * Сауришија („гуштер“; вклучува Теропода и Сауроподоморфа)
- * † Херерасауруди (рани двоножни месојади)
- * Теропода (сите двоножни; повеќето биле месојади)

- * † Целофизоиди (мали, рани тероподи; вклучува Целофисис и блиски роднини)
- * † Дилофосауриди (месојадни тероподи)
- * † Цератосаурија (важни грабливци во доцната креда на јужниот континент)
- * Тетанури („вкочанети опашки“; вклучува повеќето тероподи)
- * † Мегалосаурусиди (рана група големи месојади вклучувајќи полуакватични спиносауриди)
- * † Карносаури (Алосаури )
- * Целуросаури (пердувести тероподи, со низа големини и ниши на телото) [55]
- * † Компсогнатиди (обични рани целуросаури со намалени предни екстремитети)
- * † Тираносауриди (Тираносаурус и блиски роднини; имале намалени предни екстремитети)
- * † Орнитомимосаури ("ној; главно без заби; месојади до можни тревојади)
- * † Алваресауриди (мали инсективатори со намалени предни екстремитети, секој од нив има по една зголемена канџа)
- * Манираптори („рачни грабнувачи“; имале долги, витки раце и прсти)
- * † Теризиносауриди (двоножни тревојади со големи рачни канџи и мали глави)
- * † Овирапторосауриди (главно без заби; нивната исхрана и начин на живот не се сигурни)
- * † Археоптерикс (мали, крилести троподи или примитивни птици)
- * † Дејнонихосаури (од мала до средна големина; птица, со карактеристичен нокт на прстите)
- * Авијали (модерни птици и изумрени роднини)
- * † Скансориоптеригиди (мали примитивни авијали со долги трети прсти)
- * † Омнивороптергиди (големи, рани кратки опашки авијалани)
- * † Конфуциусорнитиди (мали авиолани без заби)
- * † Енанциорнисови птици (примитивно живеалиште на дрвја)
- * Еурнитиди (напредни летачки птици)
- * † Јанорнитиформи (птици со заби)
- * † Хесперорнити (специјализирани птици за водно нуркање)
- * Авес (модерни птици со клунови и нивни изумрени роднини)

- * † Сауроподоморфи (тревојади со мали глави, долги вратови, долги опашки)
- * † Гвибасауриди (мали, примитивни, сештојади сауроподоморфи)
- * † Платеосауриди (примитивни, строго двоножни „прозауроподи“)
- * † Риохасауриди (мали, примитивни сауроподоморфи)
- * † Масоспондилиди (мали, примитивни сауроподоморфи)
- * † Сауроподи (многу голем и тежок, обично долг над 15 метри; четириножник)
- * † Вулканодонтиди (примитивни сауроподи)
- * † Еусауроподи („вистински сауроподи“)
- * † Цетиосауриди („китови влекачи“)
- * † Туријасауриди (европска група на сауроподи од периодот на јура и креда )
- * † Неосауроподи („нови сауроподи“)
- * † Диплодокоиди (черепи и опашки издолжени; забите обично се тесни и слични на молив)
- * † Макронарија (заби во форма на лажица или молив)
- * † Брахиосауроиди (макронари со долг врат)
- * † Титаносауриди
- * † Сауроподоморфи (тревојади со мали глави, долги вратови, долги опашки)

- * † Орнитишија („птичји кошули“; разновидни двоножни и четириножни тревојади)
- * † Хетеродонтосауриди (мали базални тревојади / сештојади со истакнати заби слични на кучиња)
- * † Тиреофори (оклопни диносауруси; претежно четириножни)
- * † Анкилосаурус
- * † Стегосаури (шилкасти плочи како примарен оклоп)
- * † Неорнитишиди („нови Орнитишијци“)
- * † Орнитоподи (различни големини;)
- * † Маргиноцефалиди (се одликува со кранијален раст)
- * † Пахицефалосаури
- * † Цератопсиди (четириножници; многумина имале и рогови)
Биологија
уредиПознавањето за диносаурусите потекнува од различни фосилни и нефосилни записи, вклучително и фосилизирани коски, измет, гастролити, пердуви, внатрешните органи и меките ткива.[56][57] Многу области преку своето проучување придонесуваат за нашето разбирање на диносаурусите, вклучувајќи ја физиката (особено биомеханика), хемијата, биологијата и науките за Земјата.[58][59]. Две теми од посебен интерес и студија се големината и однесувањето на диносаурусите.[60].
Големина
уреди
Досегашните докази сугерираат дека просечната големина на диносаурусот варирала во зависност од периодот во кој живееле (тријас, рана јура, доцна јура и креда)[34]. Грабливите тероподски диносауруси, кои ги окупирале повеќето копнени ниши на месојади за време на мезозоикот, најчесто спаѓаат во категоријата од 100 до 1000 кг кога се подредуваат според проценетата тежина во категории засновани на редоследот на големината, додека неодамнешните грабливи месојадници цицачи достигнуваат од 10 до 100 кг[61]. Режимот на телесните маси на мезозоичните диносауруси е помеѓу 1 до 10 метрички тони[62]. Ова нагло се спротивставува на просечната големина на кенозоичките цицачи, проценувана од Националниот музеј на природна историја со околу 2 до 5 кг тежина[63].
Сауроподите биле најголемите и најтешките диносауруси. За поголемиот дел од периодот на диносаурусите, најмалите сауроподи биле поголеми од било што друго во нивното живеалиште, а најголемите биле поредок по големина од било што друго што тогаш одело по Земјата. Џиновските праисториски цицачи како што е Парацератериумот (најголемиот копнен цицач) биле џуџести во споредба на огромните сауроподи и единствено современите китови им се приближуваат или ги надминуваат во големина[64]. Постојат неколку предложени предности за џиновската големина на сауроподите, вклучувајќи заштита од грабливост, намалување на употребата на енергија и долговечност, но можеби најважната предност била диетална. Големите животни се поефикасни во варењето на храната отколку малите животни, бидејќи храната поминува повеќе време во нивниот дигестивен систем. Ова исто така им дозволува да се хранат со храна со помала хранлива вредност од помалите животни. Остатоците од Сауроподите најчесто се наоѓаат во карпести формации толкувани како суви или сезонско суви, а можноста за јадење големи количини со ниски хранливи материи била поволна во такви средини[65].
Најголем и најмал
уреди
_adult_male_in_flight-cropped.jpg)
Научниците веројатно никогаш нема да бидат сигурни за најголемите и најмалите диносауруси што некогаш постоеле. Тоа е затоа што само мал процент на животни некогаш биле фосилизирани и повеќето од нив остануваат закопани на земјата. Неколку примероци што се обновуваат се комплетни скелети, а впечатоците за кожата и другите меки ткива се ретки. Обновата на комплетен скелет со споредување на големината и морфологијата на коските со оние од слични, попознати видови е неточна уметност, а реконструкцијата на мускулите и другите органи на животно е, во најдобар случај, процес на образована претпоставка[66].
Највисокиот и најтешкиот диносаурус познат од добрите скелети е Жирафатитан (претходно класифициран како вид на Брахиосаурус). Неговите остатоци биле откриени во Танзанија помеѓу 1907 и 1912 година. Коските од неколку слични поединци биле вметнати во скелетот, кој сега е монтиран и изложен во Природонаучниот музеј во Берлин[67]; Овој вид бил висок 12 метри и долг од 21,8 до 22,5 метри[68][69] и претставувало животно кое тежело од 30.000 до 60.000 килограми. Најдолгиот целосен диносаурус е Диплодокус долг 27 метри, откриен во Вајоминг во Соединетите Американски Држави и прикажан во Музејот за природна историја во Питсбург[70]. Најдолгиот диносаурус познат по добар фосилен материјал е „Патаготитан“, чиј скелет се наоѓа во Американскиот музеј за природна историја во Њујорк и има должина од 37 метри. Музејот на општината Кармен Фунес во Аргентина, има реконструиран монтиран скелет на Аргентиносаурус долг 39,7 метри[71].
Постоеле и поголеми диносауруси, но знаењето за нив се заснова целосно на мал број фрагментарни фосили. Повеќето од најголемите тревојади примероци што биле заведени биле откриени во 1970-тите или подоцна, и го вклучуваат масивниот Аргентиносаурус, кој можеби тежел 800.00 до 100.000 килограми и достигнувал должина од 30 до 40 метри. Едни од најдолгите диносауруси биле Диплодокус со должина од 33,5 метри[65], (поранешен Сеизмосаурус), Суперсаурус долг 33 до 34 метри[72] и Патаготитан со должина од 37 метри. Највисокиот, Сауропосејдон бил висок 18 метри и можел да достигне прозорец на шести кат. Најтешкиот и најдолгиот диносаурус можеби бил Марапунисаурус, познат само од сега изгубениот делумен рбетен нервен лак опишан во 1878 година. Екстраполатирајќи од илустрацијата на оваа коска, животното можеби било долго 58 метри и да имало тежина од 122.400 кг. Сепак, бидејќи не се пронајдени други докази на сауроподи со оваа големина, а откривачот, Едвард Дринкер Коуп, претходно правел типографски грешки, веројатно станува збор за екстремно преценување[73].
Најголемиот месојаден диносаурус бил Спиносаурус, достигнувајќи должина од 12,6 до 18 метри и тежина од 7 до 20,9 метрички тони[74][75]. Другите големи месојадни тероподи биле Гиганотосаурус, Кархародонтосаурус и Тираносаурус[75] . Теризиносаурус и Деинохеирус биле меѓу највисоките од троподите. Најголемиот орнитишки диносаурус бил веројатно хадросауридот Шантунгосаурус кој бил со димензии 16,6 метри[76]. Најголемите единки можеби тежеле дури 16 метрички тони[77].
Најмалиот познат диносаурус е пчелесто колибри, со должина од само 5 сантиметри и тежина од околу 1,8 грамаови. Најмалите познати не-авијалски диносауруси имале големина на гулаби и биле тероподи кои биле најтесно поврзани со птиците[78]. На пример, Анхиорнис е моментално најмалиот не-авијални диносаурус опишан од примерок, со проценета тежина од 110 грама и вкупна должина на скелетот од 34 сантиметри[79]. Најмалите тревојади не-авијални диносауруси биле Микроцератус и Вананосаурус, со должина од околу 60 сантиметри[80][81].
Однесување
уреди
Голем дел од современите птици се доста социјални, и честопати се среќаваат да живеат во стада. Постои општа согласност дека некои однесувања кои се вообичаени кај птиците, како и кај крокодилите (најблиските роднини на птиците), биле исто така чести меѓу изумрените групи на диносауруси. Толкувањата на однесувањето кај фосилните видови генерално се засноваат на поза на скелети и нивно живеалиште, компјутерски симулации на нивната биомеханика и споредби со современи животни во слични еколошки ниши[58].
Првиот потенцијален доказ дека тие живееле во стадо, како широко распространето однесување и заедничко за многу групи диносауруси покрај птиците, било откритието на 31 игуандон во 1878 година во Бернисарт, Белгија, за кои тогаш се сметало дека пропаднале во дупка поради голема поплава и се удавиле[82]. Други места за масовна смрт биле откриени последователно. Оние, заедно со повеќе патеки, сугерираат дека ова однесување било вообичаено кај многу рани видови диносауруси. Откриените патеки од стотици или дури илјадници тревојади укажуваат на тоа дека хадросауридите можеби започнале да живеат во големи стада, како американскиот бизон или африканскиот спрингбок. Остатоците укажуваат дека овие животни патувале во групи составени од неколку различни видови, барем во Оксфордшир, Англија[83], иако нема докази за специфични структури на стадо[84]. Конгрегацијата во стада можеби еволуирала поради одбрана, за миграциски цели или за да обезбеди заштита на младите. Постојат докази дека многу видови на бавно растечки диносауруси, вклучувајќи разни тероподи, сауроподи, анкилосаури, орнитоподи и цератопси, како стадо сочинувале малолетните диносауруси. Еден пример е локалитет во Внатрешна Монголија, кој дал остатоци од над 20 синорнитомимуси, кои се од една до седум години на возраст. Овој склоп се толкува како социјална група која била заробена во кал[85]. Оваа особина на диносаурусите се проширила и кај месојадните тероподи како ловци на глутница кои работат заедно за да соборат голем плен[86][87]. Сепак, овој животен стил е невообичаен кај современите птици, крокодили и други влекачи, а тафонските докази кои сугерираат на лов на глутница слични на цицачи во такви троподи како Деинонихус и Алосаурус, исто така, може да се толкуваат како резултат од фатални спорови меѓу исхранувањето на животните, како што се гледа кај многу современи грабливци[88]

Грбовите на некои диносауруси, како што се маргиноцефали, тероподи и ламбеосаурини, можеби биле премногу кревки за да се користат за активна одбрана, и затоа најверојатно биле користени за сексуални или агресивни прикази, иако малку се знае за парењето на диносаурусите и територијализмот. Раните на главата од каснувања сугерираат дека троподите биле вклучени во активни агресивни конфронтации[89].
Од гледна точка на однесувањето, еден од највредните фосили на диносауруси бил откриен во пустината Гоби во 1971 година. Фосилот вклучувал велоцираптор кој напаѓал протоцератоп[90], обезбедувајќи докази дека диносаурусите навистина се нападнале едни со други. Дополнителен доказ за напад на жив плен е делумно заздравената опашка на едмонтосаурус; опашката е оштетена на таков начин што покажува дека животното било каснато од Тираносаурус, но преживеало[91]. Канибализмот кај некои видови диносауруси бил потврден со траги од заби пронајдени во Мадагаскар во 2003 година, во кои учествувал тероподот Мајунгасаурус[92].
Споредби помеѓу склеротишките (Sclerotic) прстени на диносаурусите и современите птици и влекачи биле користени за да се заклучи дневната активност на диносаурусите. Иако се претпоставува дека повеќето диносауруси биле активни во текот на денот, овие споредби покажале дека малите грабливи диносауруси како што се дромеосауриди, јураванатор и мегапносаурус биле веројатно ноќни грабливци. Големи и средни тревојади и сештојади диносауруси, како што се цератопси, сауроподоморфи, хадросауриди, орнитомиосаури, можеби биле катемерни, биле активни во кратки интервали во текот на денот, иако се сметало дека малиот Агилисаурус е дневен[93].
Врз основа на сегашните фосилни докази од диносаурусите, како што е Ориктодром, се смета дека делумно воделе фосоријален (дупчен) начин на живот[94]. Многу современи птици се арбореални (можат да се искачат на дрво), а тоа важело и за многу мезозоски птици, особено за енанциорнисовите птици[95]. Додека некои рани видови слични на птици можеби веќе биле арбореални (вклучително и дромеосауридите како што е Микрораптор[96]), повеќето неавијалани диносауруси се смета дека се потпирале на копненото движење. Доброто разбирање за тоа како се движеле диносаурусите на земја е клучно за моделите на однесување на диносаурусите; науката за биомеханика обезбедила значаен увид во оваа област. На пример, студиите за силите што ги вршат мускулите и гравитацијата врз скелетната структура на диносаурусите, дале податок за тоа колку брзо би можеле да трчаат диносаурусите[58], дали диплодоцидите можат да создадат звучни бум преку крцкање на опашката како камшик[97], и дали сауроподите можат да лебдат[98].
Комуникација
уреди
Познато е дека современите птици комуницираат користејќи визуелни и аудитивни сигнали, а широката разновидност на структурите на визуелниот приказ меѓу фосилните диносаурусни групи, како што се рогови, едра и пердуви итн., сугерираат дека визуелната комуникација отсекогаш била важна во биологијата на диносаурусите[99]. Реконструкција на бојата на перјата на анхиорнисот, укажува на важноста на бојата во визуелната комуникација кај не-птичјите диносауруси[100]. Еволуцијата на вокализацијата на диносаурусите е помалку сигурна. Палеонтологот Фил Сентер сугерира дека не-птичјите диносауруси најмногу се потпирале на визуелноста и евентуално невокалните акустични звуци како свирење, мелење со вилицата или плескање, удирање со крилјата. Тој наведува дека веројатно не биле во можност да вокализираат бидејќи нивните најблиски роднини, крокодилите и птиците, користат различни средства за да вокализираат, првите преку гркланот, а вторите преку уникатниот сиринкс, што укажува дека тие еволуирале независно, а нивниот заеднички предок бил нем[99].
Најраните остатоци од сиринкс, кој има доволно минерална содржина за фосилизација, е пронајдено во примерок од вегавис, датиран пред 69-66 милиони години, а овој орган веројатно не постоел кај нептичјите диносауруси. Сепак, за разлика од Сентер, истражувачите сугерираат дека диносаурусите можат да вокализираат и дека гласилниот систем на птиците засновани на сиринкс еволуирал од ларинксот, наместо двата система да се развиваат независно[101]. Студија од 2016 година сугерира дека диносаурусите создавале вокализирање преку затворена уста, како што е гугање, што се јавува и кај крокодилите и кај птиците, како и кај други влекачи. Ваквите вокализации еволуирале независно кај постојните архосауруси многу пати, по зголемувањето на големината на телото[102]. Се претпоставува дека сиринксот на ламбеосаурусот и носните комори на анкилосауриди функционираат во вокална резонанца[103][104], иако Сентер наведува дека присуството на комори за резонанца во некои диносауруси не е доказ за вокализација, бидејќи современите змии имаат такви комори[99].
Репродуктивна биологија
уреди
Сите диносауруси снесувале амнионски јајца со тврди лушпи направени претежно од калциум карбонат[105]. Јајцата на диносаурусите обично се поставувале во гнездо. Повеќето видови создавале гнезда во вид на чаши, куполи, чинии, или пак ги оставале во некои насипи или дупки[106]. Некои видови на модерни птици немаат гнезда; некои ги положуваат своите јајца на голи карпи, а машките царски пингвини ги чуваат јајцата помеѓу телото и нозете. Примитивните птици и многу не-авијалански диносауруси често положуваат јајца во комуналните гнезда, а мажјаците првенствено ги инкубираат јајцата. Додека современите птици имаат само еден функционален јајцевод и лежат на едно јајце одеднаш, попримитивните птици и диносаурусите имале два јајцеводи, како крокодилите. Кај некои диносауруси, како што е тродонот, положувале пар јајца на секој еден или два дена[107].
При поставување јајца, на женките им растело посебен вид коска помеѓу тврдата надворешна коска и сржта на нивните екстремитети. Оваа медуларна коска, богата со калциум, се користила за производство на лушпи од јајца. Откритието на одликите во скелетот на Тираносаурус Рекс обезбедило докази за медуларна коска во изумрени диносауруси и, за прв пат, им овозможики на палеонтолозите да го утврдат полот на примерок од фосилен диносаурус. Понатамошни истражувања откриле медуларна коска во карносаурусот Алосаурус и орнитоподот Тенонтосаурус. Бидејќи линијата на диносауруси, која ги вклучува Алосауруси Тираносаурус, се разидувала од линијата што довела до Тенонтосаурус многу рано во еволуцијата на диносаурусите, ова сугерира дека производството на медуларно ткиво е општа одлика на сите диносауруси[108].

Друга широко распространета одлика кај современите птици е родителска грижа за младите по излегувањето од јајцето. Откривањето на Хорнер во 1978 година за гнездење на Мајасаура („добра мајка гуштер“) во Монтана покажало дека родителската грижа продолжила долго по раѓањето меѓу орнитоподи[109]. Еден примерок од монголскиот овирапторид Цитипати кој бил откриен во 1993 година[110], покажал дека тие започнале да користат изолационен слој пердуви за да ги задржат јајцата топли. Ембрион на диносаурус бил пронајден без заби, што укажува на тоа дека била потребна родителска грижа за да се хранат младите диносауруси[111]. Фосилните патеки исто така го потврдиле однесувањето на родителите меѓу орнитоподите од островот Скај во северозападна Шкотска[112]. Сепак, има многу докази за претсоцијалност или суперпрецијалност меѓу многу видови диносауруси, особено тероподите. На пример, докажано е дека птиците што не се орнитуроморфи имаат бавни стапки на раст, однесување за закопување на јајца и можност за летање наскоро по раѓањето[113][114][115][116]. И Тираносаурусот Рекс и тродонот прикажуваат малолетници со јасна суперпрецијалност и веројатно зафаќаат различни еколошки ниши од возрасните.
Физиологија
уредиБидејќи и современите крокодили и птиците имаат четири комори на срцето (иако изменети кај крокодилите), се смета дека дека ова е одлика што ја делат сите архосауруси, вклучително и сите диносауруси.[117]. Додека сите современи птици имаат висок метаболизам и се „топлокрвни“ (ендотермични), од 60-тите години на минатиот век се води силна дебата во врска со тоа колку се протегала оваа особина назад во лозата на диносаурусите. Научниците не се согласуваат околу тоа дали нетичјите диносауруси биле ендотермични, ектотермични или некаква комбинација од двете[118].
Откако биле откриени нептичји диносауруси, палеонтолозите најпрвин тврделе дека тие се ектотермични. Оваа претпоставена „ладнокрвност“ се користела за да имплицира дека античките диносауруси биле релативно бавни, со слаби организми, иако многу современи влекачи се брзи и покрај тоа што се потпираат на надворешни извори на топлина за регулирање на нивната телесна температура. Идејата за диносаурусите како ектотермични останала сè додека Роберт Т. „Боб“ Бакер, раниот поборник на ендотермијата на диносаурусите, не објавил влијателен труд на оваа тема во 1968 година[119][120].
Современите докази покажуваат дека некои нептичји диносауруси напредувале во поладна умерена клима и дека некои рани видови сигурно ја регулирале телесната температура со внатрешни биолошки средства (потпомогнато од најголемиот дел од животните кај големи видови и пердуви или други прекривки на телото кај помали видови). Доказите за ендотермија кај мезозоиските диносауруси вклучуваат откривање на поларни диносауруси во Австралија и Антарктикот, како и анализа на структурите на крвните садови во фосилните коски, карактеристични за ендотермите. Продолжува научната дебата во врска со специфичните начини на кои еволуирала регулацијата на температурата на диносаурусите[121][122].

Кај сауриските диносауруси, повисоките метаболизами биле поддржани од еволуцијата на птичјиот респираторен систем, кој се одликува со широк систем на воздушни вреќи кои ги прошируваат белите дробови и напаѓаат многу коски во скелетот, правејќи ги шупливи. Раните респираторни системи во птичји стил со воздушни вреќи можеби биле способни да одржат поголеми нивоа на активност од оние на цицачите со слична големина и градба. Покрај обезбедувањето многу ефикасно снабдување со кислород, брзиот проток на воздух би бил ефикасен механизам за ладење, што е од суштинско значење за животни кои се активни, но премногу големи за да се ослободат од целата вишок на топлина преку нивната кожа[123].
Како и другите влекачи, диносаурусите се првенствено урикотелни, односно нивните бубрези извлекуваат азотни отпадоци од крвотокот и ги излачуваат како урична или мочна киселина наместо уреа или амонијак преку уретерите во цревата. Кај повеќето живи видови, уричната киселина се излачува заедно со изметот како полусилиден отпад[124][125][126]. Сепак, барем некои современи птици (како што се колибрите) може да бидат факултативно амонотелни, излачувајќи ги повеќето азотни отпадоци како амонијак[127]. Овој материјал, како и излезот на цревата, излегува од клоаката[128][129]. Покрај тоа, многу видови повторно ги регетираат пелетите, а фосилните пелети кои можеби потекнувале од диносаурусите се познати уште од дамнешно време во периодот креда.
Потекло на птици
уредиМожноста дека диносаурусите биле предци на птиците, за првпат била предложена во 1868 година од Томас Хенри Хаксли[130]. По работата на Герхард Хајлман на почетокот на XX век, теоријата за птиците како потомци на диносауруси била напуштена во корист на идејата тие да бидат потомци на генерализирани текодонти, со доказ за наводниот недостаток на клучна коска во диносаурусите[131]. Меѓутоа, како што покажале подоцнежните откритија, клучната коска всушност не била отсутна[10]. Тие биле пронајдени уште во 1924 година во Овираптор, но погрешно идентификувани[132]. Во 70-тите години на минатиот век, Џон Остром ја оживеал теоријата на диносаурус-птица[133], која добила интензитет во следните децении со доаѓањето на кладистичката анализа[134], и големиот пораст на откривањето на мали троподи и рани птици. Особено се забележуваат фосилите на формацијата Јиксијан, каде се пронајдени различни троподи и рани птици, често со пердуви од некаков вид[10][55][135]. Птиците имаат над сто различни анатомски одлики со диносаурусите тероподи, за кои денес се прифаќа дека биле нивни најблиски антички роднини[136]. Малцинство научници предложиле и други еволутивни патеки, вклучително и ревидирани верзии на предлогот на базалниот архосаурус на Хајлман, или дека манирапторските троподи се предци на птиците, но самите не се диносауруси, само што се конвергираат со диносаурусите[137].
Пердуви
уреди
Пердувите се една од најпрепознатливите одлики на современите птици, и особина што ја споделувале сите други групи на диносауруси. Врз основа на сегашната распространетост на фосилни докази, се смета дека пердувите биле одлики на диносаурусите, иако оваа одлика селективно се изгубила кај некои видови[138]. Директни фосилни докази за пердуви или структури слични на пердуви биле откриени во разновидна низа видови во многу групи нептичји диносауруси[55], и меѓу сауридите и орнитишите. Едноставни, разгранети, структури слични на пердуви се познати од хетеродонтосауридите, примитивни неорнитишиди[139] и тероподи[140], и примитивни цератопсиди. Докази за вистински, безпердувини, но сепак слични на пердувите за летање на современите птици, биле пронајдени единствено во подгрупата на тероподите Манираптора, која вклучува оварапторосаури, тродондиди, дромеосауриди и птици[10][141]. Структури слични на пердуви познати како пикнофибри се пронајдени и во птеросаурусите[142], што укажува на можноста дека филаментите слични на пердуви биле чести во лозата на птиците и се развиле пред појавата на самите диносауруси[138]. Археоптериксот бил првиот пронајден фосил што открил потенцијална врска помеѓу диносаурусите и птиците. Се смета за преоден фосил, со тоа што прикажува одлики на обете групи. Откриено е само две години по објавената на Чарлс Дарвин „За потеклото на видовите“ (1859), и неговото откритие ја поттикнало дебатата меѓу заговорниците на еволутивната биологија и креационизмот. Оваа рана птица е толку слична на диносауруси, што нема јасен впечаток на пердуви во околната карпа, и барем еден примерок бил погрешно наречен Компсогнат[143]. Од 90-тите години на минатиот век, пронајдени биле бројни дополнителни перјасти диносауруси, обезбедувајќи уште посилни докази за тесната врска помеѓу диносаурусите и современите птици[144] . Повеќето од овие примероци биле откриени на формацијата Јиксијан, североисточна Кина, која била дел од островскиот континент за време на мезозоикот. Иако биле пронајдени пердуви на само неколку локации, можно е да се наоѓаат и нептичји диносауруси на друго место во светот. Недостатокот на широко распространети докази за фосили за перјасти нептичји диносауруси може да биде затоа што деликатните одлики како кожата и пердувите не се зачувуваат често со фосилизација и затоа отсуствуваат од фосилните записи[145].
Описот на пердувести диносауруси не останал без контроверзии. Можеби најгласните критичари биле Алан Федучија и Теагартен Лингам-Солиар, кои предложиле дека некои наводни фосили како пердуви се резултат на распаѓање на колагенски влакна што ја поткрепуваат кожата на диносаурусите[146][147][148], и дека диносаурусите од типот манирапторан со пердуви всушност не биле диносауруси, туку конвергентни со диносаурусите[137][147]. Сепак, нивните ставови во најголем дел не биле прифатени од другите истражувачи, до тој степен што била доведена во прашање научната природа на предлозите на Федучија[149].
Во 2016 година било објавено дека е пронајдена опашка диносаурус со пердуви затворена во килибар. Фосилот е стар околу 99 милиони години[55][150][151].
Скелет
уредиБидејќи пердувите често се поврзуваат со птиците, овие диносауруси често се промовираат како алката што недостасува помеѓу птиците и диносаурусите. Сепак, повеќекратните одлики на скелетот, исто така споделени од двете групи, претставуваат уште една важна линија на докази за палеонтолозите. Областите на скелетот со важни сличности вклучуваат врат, препонска коска, зглоб, рачниот појас, фуркула. Споредбата на скелетите на птиците и диносаурусите преку кладистичка анализа го зацврстува случајот за нивната врската[152].
Мека анатомија
уреди
Големите диносауруси кои јаделе месо имале комплексен систем на воздушни вреќички слични на оние пронајдени кај современите птици, според истрагата спроведена во 2005 година, предводена од Патрик М. О'Конор. Белите дробови на тероподните диносауруси (месојади кои оделе на две нозе и имале стапала како птица) најверојатно пумпале воздух во шупливи вреќи во своите скелети, како што е случајот со птиците. „Она што некогаш формално се сметаше за уникатно за птиците, беше присутно во одредена форма кај предците на птиците“, рекол О’Конор[153][154]. Во 2008 година, научниците го опишале Аеростеон, чиј скелет обезбедува најсилен доказ до сега за диносаурус со систем за дишење сличен на птица. Компјутерската томографија на фосилните коски на Аеростеон открила докази за постоење воздушни вреќи во телесната празнина на животното[155][156].
Докази за однесувањето
уредиФосилите на троодонтите Меј Лонг и Синорнитоидес демонстрираат дека некои диносауруси спиеле со напикани глави под рацете[157]. Ова однесување, кое можеби помогнало да се загрева главата, е карактеристично и за современите птици. Неколку примероци како deinonychosaur и oviraptorosaur се исто така пронајдени зачувани на врвот на нивните гнезда, и веројатно се размножувале на начин како птицата[158]. Односот помеѓу волуменот на јајцето и телесната маса на возрасните меѓу овие диносауруси сугерира дека јајцата првенствено биле размножувани од мажјаците и дека младите биле многу предсоцијални, слично на многу современи птици што живеат на земјата[159][160].
Исчезнување на главните групи
уредиОткритието дека птиците се вид на диносаурус покажало дека диносаурусите воопшто не се исчезнати како што се смета вообичаено[161]. Сепак, сите нептичји диносауруси, за кои се проценува дека биле 628-1078 видови [162][163], како и многу групи птици ненадејно исчезнале пред околу 66 милиони години. Се претпоставува дека затоа што малите цицачи, лушпарите и птиците ги окупирале еколошките ниши, прилагодени на малата големина на телото, нептичји диносауруси никогаш не еволуирале во разновидна фауна на мали телесни видови, што довело до нивно пропаѓање кога биле погодени тереподите со тешките тетраподи со настанот за масовно истребување[164]. Во тоа време исчезнале и многу други групи животни, вклучувајќи амонити (мекотели слични на наутилус), мозаури, плесиосаури, птеросаури и многу групи цицачи[20]. Значајно е што инсектите не претрпеле забележлива загуба на популација, што ги оставило на располагање како храна за другите преживеани. Ова масовно истребување е познато како настан на изумирање креда-палеоген. Природата на настанот што го предизвикало ова масовно истребување е опширно проучувана од 1970-тите; во моментов, неколку сродни теории се поддржани од палеонтолозите. Иако консензусот е дека настанот со удар бил основната причина за истребување на диносаурусите, некои научници наведуваат други можни причини или ја поддржуваат идејата дека промената на неколку фактори е одговорен за ненадејното исчезнување на диносаурусите од фосилните записи[20][165][166].
Кратер Чикшулуб
уреди
Хипотезата за удар на астероид од Валтер Алварез и неговите колеги, привлекла на големо внимание во 1980 година, и го поврзува настанот на истребување на крајот од периодот креда со ударот на болид пред околу 66 милиони години[167]. Алварез предложил дека ненадејното зголемување на нивото на иридиум, забележано низ целиот свет во карпестиот слој, претставува директен доказ за влијанието[168]. Најголемиот дел од доказите денес сугерираат дека болидот широк 5 до 15 километри паднал во близина на полуостровот Јукатан (во југоисточно Мексико), создавајќи го кратерот Чикшулуб на околу 180 километри и активирајќи го масовното истребување.
Научниците не се сигурни дали диносаурусите напредувале или опаѓале пред настанот со ударот. Некои научници предлагаат дека ударот на метеоритот предизвикал долг и неприроден пад на атмосферската температура на Земјата, додека други тврдат дека наместо тоа би се создал необичен топлотен бран. Консензус меѓу научниците кои ја поддржуваат оваа хипотеза е дека влијанието предизвикало истребување директно (со топлината од ударот на метеоритот) и индиректно (преку целосното ладење кога материјалот исфрлен од ударниот кратер го рефлектирал топлинското зрачење од сонцето)[169]. Иако брзината на истребувањето не може да се заклучи само од записите на фосилите, различните модели сугерираат дека истребувањето било исклучително брзо, со часови отколку со години[170].
Во 2019 година, научниците кои вршеле дупчења во дното на Мексико извлекле уникатен геолошки запис за, според нив, денот кога астероидот со големина на град се распрснал на планетата[171].
Деканиски трапи
уредиПред 2000 година, аргументите дека базалтите од поплавите во Деканиските трапи предизвикале истребување обично биле поврзани со ставот дека истребувањето се одвивало постепено, бидејќи се сметало дека настаните со поплавувањето на базалт започнале пред околу 68 милиони години и траеле повеќе од 2 милиони години. Сепак, постојат докази дека две третини од Деканиските трапи биле создадени за само 1 милиони години пред околу 66 милиони години и затоа овие ерупции би предизвикале прилично брзо истребување, можеби за период од илјадници години.
Деканиските трапи во Индија би можело да предизвикаат истребување преку неколку механизми, вклучувајќи ослободување во воздухот на прашина и сулфурни аеросоли, што може да ја блокира сончевата светлина и со тоа да ја намали фотосинтезата кај растенијата. Покрај тоа, вулканизмот можеби испуштил емисии на јаглерод диоксид, што би го зголемило ефектот на стаклена градина кога прашината и аеросолите ќе се исчистат од атмосферата. Пред масовното истребување на диносаурусите, ослободувањето на вулкански гасови за време на формирањето на Деканиските трапи „придонесе за очигледно масовно глобално затоплување. Некои податоци укажуваат на просечен пораст на температурата од 8 степени во последните половина милион години пред случувањата кај Чикшулуб.“[172]
Во годините кога хипотезата на Деканиските трапи била поврзана со побавно истребување, Луис Алварез (кој починал во 1988 година) одговорил дека палеонтолозите биле заведени од ретките податоци. Додека неговото тврдење првично не било добро прифатено, подоцна интензивните теренски студии на фосилните отатоци му дале тежина на неговото тврдење. На крајот, повеќето палеонтолози почнале да ја прифаќаат идејата дека масовните изумирања на крајот од периодот креда во голема мера или барем делумно се должат на масивното влијание на Земјата. Сепак, дури и Валтер Алварез признал дека имало и други големи промени на Земјата дури и пред ударот, како што е падот на нивото на морето и масивните вулкански ерупции[173].
Можно преживување
уредиОстатоци од нептичји диносауруси повремено се наоѓаат над границата креда-палеоген. Во 2000 година, палеонтологот Спенсер Г. Лукас пријавил откривање на единствен десен ивер на хадросаур во басенот Сан Хуан, Ново Мексико и го опишал како доказ за палеоценски диносаурус. Формацијата во која е откриена коската е датирана во раната епоха на палеоценот, пред приближно 64,5 милиони години[174]. Доколку коската не се депонирала повторно во тој слој со акција на атмосферски влијанија, тоа ќе даде доказ дека некои популации на диносауруси можеби преживеале најмалку половина милион години во кенозоикот[175]. Други докази вклучуваат наоѓање остатоци од диносауруси во формација Хел-Крек до 1,3 метри над границата креда-палеоген, што претставува 40000 години поминато време[176] . Слични извештаи пристигнале и од други делови на светот, вклучувајќи ја и Кина. Сепак, многу научници ги отфрлиле наводните диносауруси. Директното датирање на самите коски го поддржуваат подоцнежниот датум, со методи на датирање од 64,8 ± 0,9 милиони години. Доколку ова е точно, присуството на неколку диносауруси во раниот палеоцен нема да ги промени основните факти за истребувањето[177].
Историски проучувања
уредиФосилите на диносаурусите се познати со милениуми, иако нивната вистинска природа не била препознаена. Кинезите ги сметале за коски од змеј и ги документирале како такви. На пример, во Хрониките на Хуајанг (265–316), било објавено откривање на коски од змеј во провинцијата Сечуан. Селаните во централна Кина одамна откриле фосилизирани „коски од змеј“ за употреба во традиционални лекови[178]. Во Европа, генерално, се верувало дека фосилите на диносаурусите се остатоци од гиганти и други библиски суштества[179].
Рани истражувања
уредиНаучните описи за тоа што денес би биле признати како коски на диносауруси, првпат се појавиле на крајот на 17 век во Англија. Дел од коска, за која денес се знае дека е бутна коска на мегалосаурус[180], била извлечена од варовнички каменолом во Корнвел, Оксфордшир, во 1676 година. Фрагментот бил испратен до Роберт Плот, професор по хемија на Оксфордскиот универзитет, кој објавил опис во неговата Природна историја на Оксфорд-Шајр (The Natural History of Oxford-shire) во 1677 година[181]. Тој правилно ја идентификувал коската како долен екстремитет на бутна коска на големо животно и препознал дека е премногу голема за да припаѓа на кој било познат вид. Затоа, заклучил дека тоа е коска на огромен човек, можеби Титан или друг вид гигант, прикажан во легендите[182][183]. Едвард Лујд, пријател на Сер Исак Њутн, ја објавил „Lithophylacii Britannici ichnographia“ (1699), првиот научен третман на она што денес би било признато како диносаурус кога опишал и именувал заб на сауропод[184][185], кој бил пронајден во Касвел, Оксфордшир[186].

Помеѓу 1815 и 1824 година, свештеникот Вилијам Бакленд, првиот читател на геологија на Оксфордскиот универзитет, собрал повеќе фосилизирани коски на мегалосаурус и станал првата личност што опишала диносаурус во научно списание[180][187]. Вториот род на диносаурусите, идентификуван игуанодон, бил откриен во 1822 година од Мери Ен Мантел - сопруга на англискиот геолог Гидеон Мантел. Гедеон Мантел препознал сличности меѓу неговите фосили и коските на модерните игуани. Своите откритија ги објавил во 1825 година[188][189].
Проучувањето за овие „големи фосилни гуштери“ наскоро предизвикало голем интерес за европските и американските научници, а во 1842 година англискиот палеонтолог Ричард Овен го измислил терминот „диносаурус“. Тој препознал дека остатоците што биле пронајдени до тогаш споделувале голем број карактеристични одлики и решил да ги претстави како посебна таксономска група. Со поддршка на принцот Алберт, сопругот на кралицата Викторија, Овен го основал Музејот на природна историја во Лондон, за да ја прикаже националната колекција на фосили од диносауруси и други биолошки и геолошки експонати.[190]
Откривање во Северна Америка
уредиВо 1858 година, Вилијам Паркер Фулке го открил првиот познат американски диносаурус, во малото гратче Хадонфилд, Њу Џерси. (Иако претходно биле пронајдени фосили, нивната природа не била правилно препознаена.) Суштеството го добило името хадросаурус. Тоа претставувало исклучително важно откритие: хадросаурусот бил еден од првите пронајдени скоро целосни скелети на диносауруси (првиот бил во 1834 година, во Мејдстоун, Англија) и станало јасно дека станува збор за двоножно суштество. Ова било револуционерно откритие бидејќи, до тој момент, повеќето научници верувале дека диносаурусите оделе на четири нозе, како и другите гуштери. Откритијата на Фулке предизвикале бран интереси кај диносаурусите во Соединетите Американски Држави, позната како диносаурусна манија[191].


.png){kind=link}

Пример за диносауруската манија било силното соперништво помеѓу Едвард Дринкер Коп и Отниел Чарлс Марш, и двајцата се тркале за да бидат први кои ќе пронајдат нови диносауруси во познатата „коскена војна“. Расправијата најверојатно настанала кога Марш јавно истакнал дека реконструкцијата на скелетот на Елазмосаурус од страна на Коп има недостатоци: Коп ненамерно ја поставил главата на она што требало да биде крајот на опашката на животното. Борбата меѓу двајцата научници траела повеќе од 30 години, завршувајќи во 1897 година кога Коп починал откако го потрошил целото свое богатство на лов на диносаурус. За жал, многу вредни примероци на диносауруси биле оштетени или уништени поради грубите методи на парот: на пример, нивните копачи често користеле динамит за да откопуваат коски. Современите палеонтолози сметаат дека ваквите методи се груби и неприфатливи, бидејќи минирањето лесно ги уништува фосилните и стратиграфските докази. И покрај нивните нерафинирани методи, придонесот на Коп и Марш во палеонтологијата бил огромен: Марш открил 86 нови видови диносауруси, а Коп открил 56, вкупно 142 нови видови. Колекцијата на Коп денеска се наоѓа во Американскиот музеј на природна историја, додека онаа на Марш е изложена во Музејот за природна историја на Пибоди на универзитетот Јеил[192].
По 1897 година, потрагата по фосили на диносауруси се проширила на секој континент, вклучително и на Антарктикот. Првиот откриен диносаурус на Антарктикот, Антарктопелта, бил пронајден на островот Џејмс Рос во 1986 година[193].
Тековните „жаришта“ на диносаурусите ја вклучуваат Јужна Америка (особено Аргентина) и Кина. Кина особено произвелае многу исклучителни примероци со пердувести диносауруси поради уникатната геологија на фосилни остатоци на диносауруси, како и античката сушна клима особено погодна за фосилизација[145].
Диносауруска преродба
уреди
Областа на истражување на диносаурусите уживала наплив на активности што започнале во 1970-тите. Делумно ова било предизвикано од откритието на Дејноних од Џон Остром, активен грабливец што можеби бил топлокрвен, за разлика од тогаш преовладувачката слика на диносаурусите како слаби и ладнокрвни[121][194][195][196][197] [197][198][199]. Палеонтологијата на ’рбетниците станала глобална наука. Големи нови откритија на диносауруси биле направени од палеонтолози кои работеле во претходно неискористени региони, вклучувајќи ги Индија, Јужна Америка, Мадагаскар, Антарктикот и најзначајно Кина[55] (добро сочуваните пердувести диносауруси во Кина дополнително ја консолидирале врската помеѓу диносаурусите и нивните живи потомци, современите птици). Широко распространетата примена на кладистика, која ригорозно ги анализира односите меѓу биолошките организми, исто така се покажала неверојатно корисна во класификацијата на диносаурусите. Кладистичката анализа, покрај другите современи техники, помага да се компензира честопати нецелосниот и фрагментарниот фосилен запис[194].
| Времеплов на значајни описи на таксономски диносауруси |
|---|
 |
Меко ткиво и ДНК
уреди
Еден од најдобрите примери на впечатоци од меко ткиво во фосилен диносаурус е откриен во Пјетрароја Платенкалк во јужна Италија. Откритието било пријавено во 1998 година и го опишало примерокот на мал, многу млад целуросаурус, Сципионикс. Фосилот вклучува делови од цревата, дебелото црево, црниот дроб, мускулите и душникот на овој незрел диносаурус[56].
Во март 2005 година во списанието „Science“, палеонтологот Мери Хигби Швајцер и нејзиниот тим објавиле откривање на флексибилен материјал кој наликува на вистинско меко ткиво во коска на ногата на Тираносаурус рекс стара 68 милиони години во формацијата Хел Крек во Монтана. По закрепнувањето, ткивото било повторно хидрирано од научниот тим[57]. Кога фосилизираната коска била третирана во текот на неколку недели за да се отстрани минералната содржина од фосилизираната празнина на коскената срцевина (процес наречен деминерализација), Швајцер нашла докази за недопрени структури како што се крвните садови, коскената матрица и сврзното ткиво (коскени влакна). Прегледот под микроскоп дополнително открил дека наводното меко ткиво на диносаурусот има задржано фини структури (микроструктури) дури и на клеточно ниво. Точната природа и состав на овој материјал и последиците од откривањето на Швајцер, сè уште не се јасни[57].
Во 2009 година, тим вклучително и Швајцер објавиле дека, користејќи уште повнимателна методологија, тие ги дуплирале своите резултати наоѓајќи слично меко ткиво во диносаурусот [[Брахилофосаурус]], пронајден во формацијата Џудит во Монтана. Ова вклучувало уште подетално ткиво, сè до сочуваните коскени клетки кои се чини дека имаат видливи остатоци од јадра и се чини дека се црвени крвни клетки. Меѓу другите материјали пронајдени во коската, бил и колагенот, како и во коската на Тираносаурусот. Видот на колаген што животното го има во своите коски варира во зависност од неговата ДНК и, во двата случаи, овој колаген бил од ист тип кој се наоѓал во современите кокошки и ноеви[200].
Екстракцијата на античка ДНК од фосили на диносауруси е пријавена во две одделни прилики[201]; по понатамошна проверка и преглед на врсниците, сепак, ниту еден од овие извештаи не може да се потврди[202]. Сепак, функционален пептид вклучен во визијата на теоретски диносаурус е изведен со употреба на аналитички методи на филогенетска реконструкција на генските низи на сродни современи видови како што се влекачи и птици[203]. Покрај тоа, неколку белковини, вклучително и хемоглобин[204], се откриени во фосили на диносауруси[205][206].
Во 2015 година, истражувачите објавиле дека наоѓаат структури слични на крвни клетки и колагенски влакна, зачувани во коскените фосили на шест примероци диносауруси од периодот на креда, стари приближно 75 милиони години[207][208].
Културни прикази
уреди

Според човечките стандарди, диносаурусите биле суштества со фантастичен изглед и честопати огромна големина. Како такви, тие ја доловиле народната фантазија и станале траен дел од човечката култура. Внесувањето на зборот „диносаурус“ на заедничкиот народен јазик ја рефлектира културната важност на животните: на англиски јазик, „диносаурус“ најчесто се користи за да се опише сè што е непрактично големо, застарено или предвидено да се истребува[209].
Ентузијазмот на јавноста за диносаурусите првпат се развил во Викторијанска Англија, каде во 1854 година, три децении откако настанале првите научни описи на диносаурусите, во лондонскиот Кристал Палас Парк биле откриени скулптури на диносауруси. Диносаурусите се покажале толку популарни што наскоро се развил силен пазар преку помали реплики. Во следните децении, изложбите на диносауруси се отвориле во парковите и музеите ширум светот, осигурувајќи дека последователните генерации ќе бидат запознаени со животните на извонреден и возбудлив начин[210]. Трајната популарност на диносаурусите, од своја страна, резултирало со значително јавно финансирање на науката за диносаурусите и честопати поттикнувало нови откритија. На пример, во САД, натпреварот меѓу музеите за внимание на јавноста довел директно до т.н. Коскени војни од 1880-тите и 1890-тите, за време на кои еден пар палеонтолози дале огромен научен придонес[211].
Популарната преокупација со диносаурусите го обезбедил нивниот изглед во литературата, филмот и другите медиуми. Почнувајќи од 1852 година со минувачко споменување во „Bleak House“ на Чарлс Дикенс[212], диносаурусите се прикажани во голем број измислени дела. Романот „Патување до центарот на Земјата“ на Жил Верн од 1864 година, книгата „Изгубениот свет“ на Сер Артур Конан Дојл од 1912 година, иконскиот филм „Кинг Конг“ од 1933 година, „Годизила“ од 1954 година и неговите многу други продолженија, најпродаваниот роман „Паркот Јура“ од Мајкл Крихтон од 1990 година и неговата филмската адаптација од 1993 година е само неколку забележителни примери на појава на диносаурус во фикцијата. Диносаурусите се сеприсутни во рекламирањето; бројни компании искористувале диносауруси во печатени или телевизиски реклами, или со цел да продаваат свои производи или со цел да ги карактеризираат своите соперници како бавни, слабо духовити или застарени[213].
Диносаурусите како тема во уметноста и во популарната култура
уредиДиносаурусите како тема во книжевноста
уреди- „Диносауруси“ - расказ на италијанскиот писател Итало Калвино.[214]
- „хај концепт и врската со диносаурусите“ - краток расказ на македонскиот писател Хигсов Бозон од 2011 година.[215]
Диносаурусите како тема во музиката
уреди- „Прошетај го диносаурусот“ (Walk the Dinosaur) - песна на американската поп-група „Воз (Нот Воз)“ (Was (Not Was)) од 1988 година.[216]
Диносаурусите како тема во филмот
уреди- „Добриот диносаурус“ (The Good Dinosaur) - американски анимиран филм од 2015 година.[217]
- „Јурски парк“ (Jurassic Park) - американски филм од 1993 година, во режија на Стивен Спилберг.
- „Загубениот свет: Јурски парк“ (The Lost World: Jurassic Park) - американски филм од 1997 година, во режија на Стивен Спилберг.
- „Јурски парк 3“ (Jurassic Park III) - американски филм од 2001 година, во режија на Џо Џонстон.
- „Јурски свет“ (Jurassic World) - американски филм од 2015 година, во режија на Колин Треворов.
Белешки
уреди- ↑ Иако диносаурусите се влекачи, ставовите за нив како ладнокрвни животни се промениле во последниве години; погледнете Диносаурус § Белешки за повеќе детали
Наводи
уреди- ↑ 1,0 1,1 Owen 1842, p.103: "The combination of such characters … will, it is presumed, be deemed sufficient ground for establishing a distinct tribe or sub-order of Saurian Reptiles, for which I would propose the name of Dinosauria*. (*Gr. δεινός, fearfully great; σαύρος, a lizard. … )
- ↑ Предлошка:Cite Merriam-Webster
- ↑ Crane, George R. (уред.). „Greek Dictionary Headword Search Results“. Perseus 4.0. Medford and Somerville, MA: Tufts University. Посетено на October 13, 2019. Lemma for 'δεινός' from Henry George Liddell, Robert Scott, A Greek-English Lexicon (1940): 'fearful, terrible'. Грешка во наводот: Неважечка ознака
<ref>; називот „LSJ“ е зададен повеќепати со различна содржина. - ↑ Farlow & Brett-Surman 1997, стр. ix–xi, Preface, "Dinosaurs: The Terrestrial Superlative" by James O. Farlow and M.K. Brett-Surman.
- ↑ Chamary, JV (September 30, 2014). „Dinosaurs, Pterosaurs And Other Saurs – Big Differences“. Forbes. Jersey City, NJ. ISSN 0015-6914. Архивирано од изворникот 2014-11-10. Посетено на 2018-10-02.
- ↑ 6,0 6,1 6,2 Weishampel, Dodson & Osmólska 2004, стр. 7–19, chpt. 1: "Origin and Relationships of Dinosauria" by Michael J. Benton.
- ↑ Olshevsky 2000
- ↑ 8,0 8,1 Langer, Max C.; Ezcurra, Martin D.; Bittencourt, Jonathas S.; Novas, Fernando E. (February 2010). „The origin and early evolution of dinosaurs“. Biological Reviews. Cambridge: Cambridge Philosophical Society. 85 (1): 65–66, 82. doi:10.1111/j.1469-185x.2009.00094.x. hdl:11336/103412. ISSN 1464-7931. PMID 19895605. S2CID 34530296.
- ↑ „Користење на дрвото за класификација“. Understanding Evolution. Беркели: Универзитет Калифорнија. Архивирано од изворникот 2019-08-31. Посетено на 2019-10-14.
- ↑ 10,0 10,1 10,2 10,3 10,4 Weishampel, Dodson & Osmólska 2004, стр. 210–231, chpt. 11: "Basal Avialae" by Kevin Padian.
- ↑ Вејд, Николас (22 март 2017). „Тресење на семејното стебло Диносаурус“. The New York Times. New York. ISSN 0362-4331. Архивирано од изворникот 2018-04-07. Посетено на 2019-10-30. „Верзија на овој напис се појавува во печатено издание на 28 март 2017 година, на страница Д6 од изданието во Њујорк со наслов: Тресење на семејното стебло на диносаурус .“
- ↑ Барон, Метју; Норман, Дејвид; Барет, Пол (22 март 2017). „Нова хипотеза за односите со диносаурусите и раната еволуција на диносаурусите“. Nature. Лондон: Nature Research. 543 (7646): 501–506. Bibcode:2017Natur.543..501B. doi:10.1038/nature21700. ISSN 0028-0836. PMID 28332513. S2CID 205254710. „Оваа податотека содржи дополнителен текст и податоци, дополнителни табели 1-3 и дополнителни препораки„: Дополнителни информации[мртва врска]
- ↑ Glut 1997, стр. 40
- ↑ Lambert & The Diagram Group 1990, стр. 288
- ↑ Farlow & Brett-Surman 1997, стр. 607–624, chpt. 39: "Major Groups of Non-Dinosaurian Vertebrates of the Mesozoic Era" by Michael Morales.
- ↑ Wang, Steve C.; Dodson, Peter (2006). „Estimating the diversity of dinosaurs“. Proc. Natl. Acad. Sci. U.S.A. Washington, D.C.: National Academy of Sciences. 103 (37): 13601–13605. Bibcode:2006PNAS..10313601W. doi:10.1073/pnas.0606028103. ISSN 0027-8424. PMC 1564218. PMID 16954187.
- ↑ Russell, Dale A. (1995). „China and the lost worlds of the dinosaurian era“. Historical Biology. Milton Park, Oxfordshire: Taylor & Francis. 10 (1): 3–12. doi:10.1080/10292389509380510. ISSN 0891-2963.
- ↑ Старфелт, Џостејн; Лиов, Ли (5 април 2016). „Колку видови диносауруси имало? Предрасуди на фосили и вистинско богатство проценето со примерок од Поасон“. Philosophical Transactions of the Royal Society B. Лондон: Royal Society. 371 (1691): 20150219. doi:10.1098/rstb.2015.0219. ISSN 0962-8436. PMC 4810813. PMID 26977060.
- ↑ Свитек, Бријан (23 март 2016). „Повеќето видови на диносауруси сè уште не се откриени“. Phenomena - A Science Salon (англиски). Вашингтон: National Geographic Society. OCLC 850948164. Архивирано од изворникот 2019-07-12. Посетено на 2019-11-07.
- ↑ 20,0 20,1 20,2 Грешка во наводот: Погрешна ознака
<ref>; нема зададено текст за наводите по имеMacLeod. - ↑ 21,0 21,1 Amiot, Romain; Buffetaut, Éric; Lécuyer, Christophe; и др. (2010). „Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods“. Geology. Boulder, CO: Geological Society of America. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1. ISSN 0091-7613.
- ↑ 22,0 22,1 22,2 Brusatte 2012, стр. 9–20, 21
- ↑ Nesbitt, Sterling J. (2011). „The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades“ (PDF). Bulletin of the American Museum of Natural History. New York: American Museum of Natural History. 2011 (352): 1–292. doi:10.1206/352.1. hdl:2246/6112. ISSN 0003-0090. S2CID 83493714. Архивирано од изворникот February 29, 2016. Посетено на October 16, 2019.
- ↑ Paul 2000, стр. 140–168, chpt. 3: "Classification and Evolution of the Dinosaur Groups" by Thomas R. Holtz Jr.
- ↑ Смит, Дејв; и др. „Диносаурија: Морфологија“. Беркели: Палентолошки универзитет на Универзитетот Калифорнија. Посетено на 2019-10-16.
- ↑ Langer, Max C.; Abdala, Fernando; Richter, Martha; Benton, Michael J. (1999). „Un dinosaure sauropodomorphe dans le Trias supérieur (Carnien) du Sud du Brésil“ [A sauropodomorph dinosaur from the Upper Triassic (Carman) of southern Brazil]. Comptes Rendus de l'Académie des Sciences, Série IIA. Amsterdam: Elsevier on behalf of the French Academy of Sciences. 329 (7): 511–517. Bibcode:1999CRASE.329..511L. doi:10.1016/S1251-8050(00)80025-7. ISSN 1251-8050.
- ↑ Nesbitt, Sterling J.; Irmis, Randall B.; Parker, William G. (2007). „A critical re-evaluation of the Late Triassic dinosaur taxa of North America“. Journal of Systematic Palaeontology. Milton Park, Oxfordshire: Taylor & Francis on behalf of the Natural History Museum, London. 5 (2): 209–243. doi:10.1017/S1477201907002040. ISSN 1477-2019. S2CID 28782207.
- ↑ This was recognized not later than 1909: Celeskey, Matt (2005). „Dr. W. J. Holland and the Sprawling Sauropods“. The Hairy Museum of Natural History. Архивирано од изворникот на June 12, 2011. Посетено на October 18, 2019.
- Holland, William J. (May 1910). „A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus Carnegiei in the Carnegie Museum“. The American Naturalist. American Society of Naturalists. 44 (521): 259–283. doi:10.1086/279138. ISSN 0003-0147. S2CID 84424110. Посетено на October 18, 2019.
- The arguments and many of the images are also presented in Desmond 1975.
- ↑ 29,0 29,1 Benton 2005
- ↑ Cowen 2005, стр. 151–175, chpt. 12: "Dinosaurs".
- ↑ Kump, Lee R.; Pavlov, Alexander; Arthur, Michael A. (2005). „Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia“. Geology. Boulder, CO: Geological Society of America. 33 (5): 397–400. Bibcode:2005Geo....33..397K. doi:10.1130/G21295.1. ISSN 0091-7613. S2CID 34821866.
- ↑ Tanner, Lawrence H.; Lucas, Spencer G.; Chapman, Mary G. (March 2004). „Assessing the record and causes of Late Triassic extinctions“ (PDF). Earth-Science Reviews. Amsterdam: Elsevier. 65 (1–2): 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. ISSN 0012-8252. Архивирано од изворникот (PDF) на October 25, 2007. Посетено на October 22, 2007.
- ↑ Алкобер, Оскар А.; Мартинес, Рикардо (19 октмври 2010). „Нов херерасаурид (Диносаурија, Сауришија) од Горно-тријашката формација Ишигуаласто на северозападна [[Аргентина]]“. ZooKeys. Софија: Pensoft Publishers (63): 55–81. doi:10.3897/zookeys.63.550. ISSN 1313-2989. PMC 3088398. PMID 21594020. Проверете ги датумските вредности во:
|date=(help); URL–wikilink conflict (help) - ↑ 34,0 34,1 Sereno, Paul C. (1999). „The Evolution of Dinosaurs“. Science. Washington, D.C.: American Association for the Advancement of Science. 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. ISSN 0036-8075. PMID 10381873. Архивирано (PDF) од изворникот January 5, 2018. Посетено на November 8, 2019.
- ↑ Sereno, Paul C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). „Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria“. Nature. London: Nature Research. 361 (6407): 64–66. Bibcode:1993Natur.361...64S. doi:10.1038/361064a0. ISSN 0028-0836. S2CID 4270484.
- ↑ Несбит, Стерлинг; Барет, Пол; Веринг, Сара; и др. (23 февруари 2012). „Најстариот диносаурус? Среднотриски диносауриформ од Танзанија“. Biology Letters. Лондон: Royal Society. 9 (1): 20120949. doi:10.1098/rsbl.2012.0949. ISSN 1744-9561. PMC 3565515. PMID 23221875.
- ↑ Лангер, Макс; Рамезани, Џахандар; Да Роса, Атила (мај 2018). „U-Pb возрасните ограничувања на диносаурусот се зголемуваат од јужен Бразил“. Gondwana Research. Амстердам: Elsevier. 57: 133–140. Bibcode:2018GondR..57..133L. doi:10.1016/j.gr.2018.01.005. ISSN 1342-937X.
- ↑ Brusatte, Stephen L.; Benton, Michael J.; Ruta, Marcello; Lloyd, Graeme T. (September 12, 2008). „Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs“ (PDF). Science. Washington, D.C.: American Association for the Advancement of Science. 321 (5895): 1485–1488. Bibcode:2008Sci...321.1485B. doi:10.1126/science.1161833. hdl:20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b. ISSN 0036-8075. PMID 18787166. S2CID 13393888. Посетено на 2019-10-22.
- ↑ Tanner, Spielmann & Lucas 2013, стр. 562–566, "The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A." by Justin A. Spielmann, Spencer G. Lucas and Adrian P. Hunt.
- ↑ Sulej, Tomasz; Niedźwiedzki, Grzegorz (January 4, 2019). „An elephant-sized Late Triassic synapsid with erect limbs“. Science. Washington, D.C.: American Association for the Advancement of Science. 363 (6422): 78–80. Bibcode:2019Sci...363...78S. doi:10.1126/science.aal4853. ISSN 0036-8075. PMID 30467179. S2CID 53716186.
- ↑ „Fossil tracks in the Alps help explain dinosaur evolution“. Science and Technology. The Economist. London. April 19, 2018. ISSN 0013-0613. Посетено на 2018-05-24.
- ↑ 42,0 42,1 42,2 42,3 Weishampel, Dodson & Osmólska 2004, стр. 627–642, chpt. 27: "Mesozoic Biogeography of Dinosauria" by Thomas R. Holtz Jr., Ralph E. Chapman, and Matthew C. Lamanna.
- ↑ 43,0 43,1 43,2 43,3 Weishampel, Dodson & Osmólska 2004, стр. 614–626, chpt. 26: "Dinosaur Paleoecology" by David E. Fastovsky and Joshua B. Smith.
- ↑ Sereno, Paul C.; Wilson, Jeffrey A.; Witmer, Lawrence M.; и др. (2007). Kemp, Tom (уред.). „Structural Extremes in a Cretaceous Dinosaur“. PLOS ONE. San Francisco, CA: PLOS. 2 (11): e1230. Bibcode:2007PLoSO...2.1230S. doi:10.1371/journal.pone.0001230. ISSN 1932-6203. PMC 2077925. PMID 18030355.
- ↑ Prasad, Vandana; Strömberg, Caroline A. E.; Alimohammadian, Habib; и др. (2005). „Dinosaur Coprolites and the Early Evolution of Grasses and Grazers“. Science. Washington, D.C.: American Association for the Advancement of Science. 310 (5751): 1170–1180. Bibcode:2005Sci...310.1177P. doi:10.1126/science.1118806. ISSN 0036-8075. PMID 16293759. S2CID 1816461.
- ↑ Weishampel, Dodson & Osmólska 2004, стр. 672–684, chpt. 30: "Dinosaur Extinction" by J. David Archibald and David E. Fastovsky.
- ↑ Dyke & Kaiser 2011, chpt. 14: "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification" by Bent E. K. Lindow. doi:10.1002/9781119990475.ch14
- ↑ Cracraft, Joel (June 21, 1968). „A Review of the Bathornithidae (Aves, Gruiformes), with Remarks on the Relationships of the Suborder Cariamae“ (PDF). American Museum Novitates. New York: American Museum of Natural History (2326): 1–46. hdl:2246/2536. ISSN 0003-0082. Посетено на 2019-10-22.
- ↑ Alvarenga, Herculano; Jones, Washington W.; Rinderknecht, Andrés (May 2010). „The youngest record of phorusrhacid birds (Aves, Phorusrhacidae) from the late Pleistocene of Uruguay“. Neues Jahrbuch für Geologie und Paläontologie. Stuttgart: E. Schweizerbart. 256 (2): 229–234. doi:10.1127/0077-7749/2010/0052. ISSN 0077-7749. Посетено на 2019-10-22.
- ↑ Mayr 2009
- ↑ Weishampel, Dodson & Osmólska 2004, стр. 196–210, chpt. 10: "Dromaeosauridae" by Peter J. Makovicky and Mark A. Norell.
- ↑ Paul 1988, стр. 248–250
- ↑ Weishampel, Dodson & Osmólska 2004, стр. 151–164, chpt. 7: "Therizinosauroidea" by James M. Clark, Teresa Maryańska, and Rinchen Barsbold.
- ↑ Taylor, Michael P.; Wedel, Mathew J. (February 12, 2013). „Why sauropods had long necks; and why giraffes have short necks“. PeerJ. Corte Madera, CA; London. 1: e36. doi:10.7717/peerj.36. ISSN 2167-8359. PMC 3628838. PMID 23638372.
- ↑ 55,0 55,1 55,2 55,3 55,4 St. Fleur, Nicholas (December 8, 2016). „That Thing With Feathers Trapped in Amber? It Was a Dinosaur Tail“. Trilobites. The New York Times. New York. ISSN 0362-4331. Архивирано од изворникот 2017-08-31. Посетено на 2016-12-08.
- ↑ 56,0 56,1 Dal Sasso, Cristiano; Signore, Marco (March 26, 1998). „Exceptional soft-tissue preservation in a theropod dinosaur from Italy“. Nature. London: Nature Research. 392 (6674): 383–387. Bibcode:1998Natur.392..383D. doi:10.1038/32884. ISSN 0028-0836. S2CID 4325093.
- ↑ 57,0 57,1 57,2 Schweitzer, Mary H.; Wittmeyer, Jennifer L.; Horner, John R.; Toporski, Jan K. (2005). „Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex“. Science. Washington, D.C.: American Association for the Advancement of Science. 307 (5717): 1952–1955. Bibcode:2005Sci...307.1952S. doi:10.1126/science.1108397. ISSN 0036-8075. PMID 15790853. S2CID 30456613.
- ↑ 58,0 58,1 58,2 Alexander, R. McNeill (2006). „Dinosaur biomechanics“. Proceedings of the Royal Society B. London: Royal Society. 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532. ISSN 0962-8452. PMC 1634776. PMID 16822743.
- ↑ Farlow, James O.; Dodson, Peter; Chinsamy, Anusuya (November 1995). „Dinosaur Biology“. Annual Review of Ecology and Systematics. Palo Alto, CA: Annual Reviews. 26: 445–471. doi:10.1146/annurev.es.26.110195.002305. ISSN 1545-2069.
- ↑ Weishampel, Dodson & Osmólska 2004
- ↑ Dodson & Gingerich 1993, стр. 167–199, "On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large carnivorous dinosaurs" by James O. Farlow.
- ↑ Peczkis, Jan (1995). „Implications of body-mass estimates for dinosaurs“. Journal of Vertebrate Paleontology. Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology. 14 (4): 520–533. doi:10.1080/02724634.1995.10011575. ISSN 0272-4634. JSTOR 4523591.
- ↑ „Dinosaur Evolution“. Department of Paleobiology. Dinosaurs. Washington, D.C.: National Museum of Natural History. 2007. Архивирано од изворникот на November 11, 2007. Посетено на November 21, 2007.
- ↑ Sander, P. Martin; Christian, Andreas; Clauss, Marcus; и др. (February 2011). „Biology of the sauropod dinosaurs: the evolution of gigantism“. Biological Reviews. Cambridge: Cambridge Philosophical Society. 86 (1): 117–155. doi:10.1111/j.1469-185X.2010.00137.x. ISSN 1464-7931. PMC 3045712. PMID 21251189.
- ↑ 65,0 65,1 Foster & Lucas 2006, стр. 131–138, "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878" by Kenneth Carpenter.
- ↑ Paul 2010
- ↑ Colbert 1971
- ↑ Mazzetta, Gerardo V.; Christiansenb, Per; Fariñaa, Richard A. (2004). „Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs“ (PDF). Historical Biology. Milton Park, Oxfordshire: Taylor & Francis. 16 (2–4): 71–83. CiteSeerX 10.1.1.694.1650. doi:10.1080/08912960410001715132. ISSN 0891-2963. S2CID 56028251.
- ↑ Janensch, Werner (1950). Translation by Gerhard Maier. „Die Skelettrekonstruktion von Brachiosaurus brancai“ [The Skeleton Reconstruction of Brachiosaurus brancai] (PDF). Palaeontographica. Stuttgart: E. Schweizerbart. Suplement VII (1. Reihe, Teil 3, Lieferung 2): 97–103. OCLC 45923346. Архивирано (PDF) од изворникот 2017-07-11. Посетено на 2019-10-24.
- ↑ Lucas, Spencer G.; Herne, Matthew C.; Hecket, Andrew B.; и др. (November 9, 2004). Reappraisal of Seismosaurus, a Late Jurassic Sauropod Dinosaur From New Mexico. 2004 Denver Annual Meeting (November 7–10, 2004). 36. Boulder, CO: Geological Society of America. стр. 422. OCLC 62334058. Paper No. 181-4. Архивирано од изворникот 2019-10-08. Посетено на 2019-10-25.
- ↑ Sellers, William Irvin.; Margetts, Lee; Coria, Rodolfo Aníbal; Manning, Phillip Lars (October 30, 2013). Carrier, David (уред.). „March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs“. PLOS ONE. San Francisco, CA: PLOS. 8 (10): e78733. Bibcode:2013PLoSO...878733S. doi:10.1371/journal.pone.0078733. ISSN 1932-6203. PMC 3864407. PMID 24348896.
- ↑ Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (October–December 2007). „Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny“. Arquivos do Museu Nacional. Rio de Janeiro: National Museum of Brazil; Federal University of Rio de Janeiro. 65 (4): 527–544. CiteSeerX 10.1.1.603.7472. ISSN 0365-4508. Посетено на October 26, 2019.
- ↑ Woodruff, D. Cary; Foster, John R. (2014). „The fragile legacy of Amphicoelias fragillimus (Dinosauria: Sauropoda; Morrison Formation - Latest Jurassic)“. Volumina Jurassica. 12 (2): 211–220. doi:10.5604/17313708.1130144 (неактивно 2024-12-01).
- ↑ Dal Sasso, Cristiano; Maganuco, Simone; Buffetaut, Éric; и др. (2005). „New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities“ (PDF). Journal of Vertebrate Paleontology. Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology. 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. ISSN 0272-4634. Архивирано од изворникот (PDF) на April 29, 2011. Посетено на May 5, 2011.
- ↑ 75,0 75,1 Therrien, François; Henderson, Donald M. (2007). „My theropod is bigger than yours … or not: estimating body size from skull length in theropods“. Journal of Vertebrate Paleontology. Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology. 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2. ISSN 0272-4634.
- ↑ Zhao, Xijin; Li, Dunjing; Han, Gang; и др. (2007). „Zhuchengosaurus maximus from Shandong Province“. Acta Geoscientia Sinica. Beijing: Chinese Academy of Geological Sciences. 28 (2): 111–122. ISSN 1006-3021.
- ↑ Weishampel, Dodson & Osmólska 2004, стр. 438–463, chpt. 20: "Hadrosauridae" by John R. Horner David B. Weishampel, and Catherine A. Forster.
- ↑ „Bee Hummingbird (Mellisuga helenae)“. Birds.com. Paley Media. Архивирано од изворникот 2015-04-03. Посетено на 2019-10-27.
- ↑ Xu, Xing; Zhao, Qi; Norell, Mark; и др. (February 2008). „A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin“. Chinese Science Bulletin. Amsterdam: Elsevier on behalf of Science in China Press. 54 (3): 430–435. doi:10.1007/s11434-009-0009-6. ISSN 1001-6538. S2CID 53445386.
- ↑ Holtz 2007
- ↑ Butler, Richard J.; Zhao, Qi (February 2009). „The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China“. Cretaceous Research. Amsterdam: Elsevier. 30 (1): 63–77. doi:10.1016/j.cretres.2008.03.002. ISSN 0195-6671.
- ↑ Yans, Johan; Dejax, Jean; Pons, Denise; и др. (January–February 2005). „Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)“ [Palaeontological and geodynamical implications of the palynological dating of the wealden facies sediments of Bernissart (Mons Basin, Belgium)]. Comptes Rendus Palevol (француски). Amsterdam: Elsevier of behalf of the French Academy of Sciences. 4 (1–2): 135–150. doi:10.1016/j.crpv.2004.12.003. ISSN 1631-0683.
- ↑ Day, Julia J.; Upchurch, Paul; Norman, David B.; и др. (2002). „Sauropod Trackways, Evolution, and Behavior“. Science. Washington, D.C.: American Association for the Advancement of Science. 296 (5573): 1659. doi:10.1126/science.1070167. ISSN 0036-8075. PMID 12040187. S2CID 36530770.
- ↑ Curry Rogers & Wilson 2005, стр. 252–284, chpt. 9: "Steps in Understanding Sauropod Biology: The Importance of Sauropods Tracks" by Joanna L. Wright.
- ↑ Varricchio, David J.; Sereno, Paul C.; Zhao, Xijin; и др. (2008). „Mud-trapped herd captures evidence of distinctive dinosaur sociality“ (PDF). Acta Palaeontologica Polonica. Warsaw: Institute of Paleobiology, Polish Academy of Sciences. 53 (4): 567–578. doi:10.4202/app.2008.0402. ISSN 0567-7920. S2CID 21736244. Архивирано (PDF) од изворникот March 30, 2019. Посетено на May 6, 2011.
- ↑ Lessem & Glut 1993, стр. 19–20, "Allosaurus"
- ↑ Maxwell, W. Desmond; Ostrom, John H. (1995). „Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations“. Journal of Vertebrate Paleontology. Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology. 15 (4): 707–712. doi:10.1080/02724634.1995.10011256. ISSN 0272-4634.
- ↑ Roach, Brian T.; Brinkman, Daniel L. (April 2007). „A Reevaluation of Cooperative Pack Hunting and Gregariousness in Deinonychus antirrhopus and Other Nonavian Theropod Dinosaurs“. Bulletin of the Peabody Museum of Natural History. New Haven, CT: Peabody Museum of Natural History. 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2. ISSN 0079-032X.
- ↑ Tanke, Darren H. (1998). „Head-biting behavior in theropod dinosaurs: paleopathological evidence“ (PDF). Gaia: Revista de Geociências. Lisbon: National Museum of Natural History and Science (15): 167–184. doi:10.7939/R34T6FJ1P. ISSN 0871-5424. Архивирано од изворникот (PDF) на February 27, 2008.
- ↑ „The Fighting Dinosaurs“. New York: American Museum of Natural History. Архивирано од изворникот на January 18, 2012. Посетено на December 5, 2007.
- ↑ Carpenter, Kenneth (1998). „Evidence of predatory behavior by theropod dinosaurs“ (PDF). Gaia: Revista de Geociências. Lisbon: National Museum of Natural History and Science. 15: 135–144. ISSN 0871-5424.
- ↑ Rogers, Raymond R.; Krause, David W.; Curry Rogers, Kristina (2007). „Cannibalism in the Madagascan dinosaur Majungatholus atopus“. Nature. London: Nature Research. 422 (6931): 515–518. Bibcode:2003Natur.422..515R. doi:10.1038/nature01532. ISSN 0028-0836. PMID 12673249. S2CID 4389583.
- ↑ Schmitz, Lars; Motani, Ryosuke (2011). „Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology“. Science. Washington, D.C.: American Association for the Advancement of Science. 332 (6030): 705–708. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. ISSN 0036-8075. PMID 21493820. S2CID 33253407.
- ↑ Varricchio, David J.; Martin, Anthony J.; Katsura, Yoshihiro (2007). „First trace and body fossil evidence of a burrowing, denning dinosaur“. Proceedings of the Royal Society B. London: Royal Society. 274 (1616): 1361–1368. doi:10.1098/rspb.2006.0443. ISSN 0962-8452. PMC 2176205. PMID 17374596.
- ↑ Chiappe & Witmer 2002
- ↑ Chatterjee, Sankar; Templin, R. Jack (2007). „Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui“ (PDF). Proc. Natl. Acad. Sci. U.S.A. Washington, D.C.: National Academy of Sciences. 104 (5): 1576–1580. Bibcode:2007PNAS..104.1576C. doi:10.1073/pnas.0609975104. ISSN 0027-8424. PMC 1780066. PMID 17242354. Архивирано (PDF) од изворникот August 18, 2019. Посетено на October 29, 2019.
- ↑ Goriely, Alain; McMillen, Tyler (2002). „Shape of a Cracking Whip“. Physical Review Letters. Ridge, NY: American Physical Society. 88 (24): 244301. Bibcode:2002PhRvL..88x4301G. doi:10.1103/PhysRevLett.88.244301. ISSN 0031-9007. PMID 12059302.
- ↑ Henderson, Donald M. (2003). „Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: a computational analysis“. Canadian Journal of Zoology. Ottawa: NRC Research Press. 81 (8): 1346–1357. doi:10.1139/z03-122. ISSN 0008-4301.
- ↑ 99,0 99,1 99,2 Senter, Phil (2008). „Voices of the past: a review of Paleozoic and Mesozoic animal sounds“. Historical Biology. Milton Park, Oxfordshire: Taylor & Francis. 20 (4): 255–287. doi:10.1080/08912960903033327. ISSN 0891-2963. S2CID 84473967.
- ↑ Li, Quanguo; Gao, Ke-Qin; Vinther, Jakob; и др. (2010). „Plumage Color Patterns of an Extinct Dinosaur“ (PDF). Science. Washington, D.C.: American Association for the Advancement of Science. 327 (5971): 1369–1372. Bibcode:2010Sci...327.1369L. doi:10.1126/science.1186290. ISSN 0036-8075. PMID 20133521. S2CID 206525132. Архивирано (PDF) од изворникот March 30, 2019. Посетено на November 7, 2019.
- ↑ Clarke, Julia A.; Chatterjee, Sankar; Zhiheng, Li; и др. (2016). „Fossil evidence of the avian vocal organ from the Mesozoic“. Nature. London: Nature Research. 538 (7626): 502–505. Bibcode:2016Natur.538..502C. doi:10.1038/nature19852. ISSN 0028-0836. PMID 27732575. S2CID 4389926.
- ↑ Riede, Tobias; Eliason, Chad M.; Miller, Edward H.; и др. (2016). „Coos, booms, and hoots: the evolution of closed-mouth vocal behavior in birds“. Evolution. Hoboken, NJ: John Wiley & Sons for the Society for the Study of Evolution. 70 (8): 1734–1746. doi:10.1111/evo.12988. ISSN 0014-3820. PMID 27345722. S2CID 11986423.
- ↑ Weishampel, David B. (Spring 1981). „Acoustic Analysis of Vocalization of Lambeosaurine Dinosaurs (Reptilia: Ornithischia)“ (PDF). Paleobiology. Bethesda, MD: Paleontological Society. 7 (2): 252–261. doi:10.1017/S0094837300004036. ISSN 0094-8373. JSTOR 2400478. Архивирано од изворникот (PDF) на October 6, 2014. Посетено на October 30, 2019.
- ↑ Miyashita, Tetsuto; Arbour, Victoria M.; Witmer, Lawrence M.; и др. (December 2011). „The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction“ (PDF). Journal of Anatomy. Hoboken, NJ: John Wiley & Sons. 219 (6): 661–675. doi:10.1111/j.1469-7580.2011.01427.x. ISSN 1469-7580. PMC 3237876. PMID 21954840. Архивирано од изворникот (PDF) на September 24, 2015. Посетено на October 30, 2019.
- ↑ Currie & Padian 1997, стр. 206, "Eggs, Eggshells, and Nests" by Konstantin E. Mikhailov.
- ↑ Hansell 2000
- ↑ Varricchio, David J.; Horner, John R.; Jackson, Frankie D. (September 19, 2002). „Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus“. Journal of Vertebrate Paleontology. Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology. 22 (3): 564–576. doi:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2. ISSN 0272-4634.
- ↑ Lee, Andrew H.; Werning, Sarah (2008). „Sexual maturity in growing dinosaurs does not fit reptilian growth models“. Proc. Natl. Acad. Sci. U.S.A. Washington, D.C.: National Academy of Sciences. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. doi:10.1073/pnas.0708903105. ISSN 0027-8424. PMC 2206579. PMID 18195356.
- ↑ Horner, John R.; Makela, Robert (1979). „Nest of juveniles provides evidence of family structure among dinosaurs“. Nature. London: Nature Research. 282 (5736): 296–298. Bibcode:1979Natur.282..296H. doi:10.1038/282296a0. ISSN 0028-0836. S2CID 4370793.
- ↑ „Discovering Dinosaur Behavior: 1960–present view“. Encyclopædia Britannica. Chicago, IL: Encyclopædia Britannica, Inc. Архивирано од изворникот на December 13, 2013. Посетено на October 30, 2019.
- ↑ Reisz, Robert R.; Scott, Diane; Sues, Hans-Dieter; и др. (2005). „Embryos of an Early Jurassic Prosauropod Dinosaur and Their Evolutionary Significance“ (PDF). Science. Washington, D.C.: American Association for the Advancement of Science. 309 (5735): 761–764. Bibcode:2005Sci...309..761R. doi:10.1126/science.1114942. ISSN 0036-8075. PMID 16051793. S2CID 37548361.
- ↑ Clark, Neil D. L.; Booth, Paul; Booth, Claire L.; и др. (2004). „Dinosaur footprints from the Duntulm Formation (Bathonian, Jurassic) of the Isle of Skye“ (PDF). Scottish Journal of Geology. London: Geological Society of London. 40 (1): 13–21. doi:10.1144/sjg40010013. ISSN 0036-9276. S2CID 128544813. Архивирано (PDF) од изворникот July 22, 2013. Посетено на December 12, 2019.
- ↑ Zhou, Zhonghe; Zhang, Fucheng (October 22, 2004). „A Precocial Avian Embryo from the Lower Cretaceous of China“. Science. Washington, D.C.: American Association for the Advancement of Science. 306 (5696): 653. doi:10.1126/science.1100000. ISSN 0036-8075. PMID 15499011. S2CID 34504916.
- ↑ Naish, Darren (May 15, 2012). „A drowned nesting colony of Late Cretaceous birds“. Tetrapod Zoology. Scientific American (blog). Stuttgart: Springer Nature. 306 (5696): 653. doi:10.1126/science.1100000. ISSN 0036-8733. PMID 15499011. S2CID 34504916. Архивирано од изворникот 2018-09-25. Посетено на 2019-11-16.
- ↑ Fernández, Mariela S.; García, Rodolfo A.; Fiorelli, Lucas; и др. (April 17, 2013). „A Large Accumulation of Avian Eggs from the Late Cretaceous of Patagonia (Argentina) Reveals a Novel Nesting Strategy in Mesozoic Birds“. PLOS ONE. San Francisco, CA: PLOS. 8 (4): e61030. Bibcode:2013PLoSO...861030F. doi:10.1371/journal.pone.0061030. ISSN 1932-6203. PMC 3629076. PMID 23613776.
- ↑ Deeming, Denis Charles; Mayr, Gerald (May 2018). „Pelvis morphology suggests that early Mesozoic birds were too heavy to contact incubate their eggs“. Journal of Evolutionary Biology. Hoboken, NJ: Wiley-Blackwell on behalf of the European Society for Evolutionary Biology. 31 (5): 701–709. doi:10.1111/jeb.13256. ISSN 1010-061X. PMID 29485191. S2CID 3588317.
- ↑ Weishampel, Dodson & Osmólska 2004, стр. 643–659, chpt. 28: "Physiology of Nonavian Dinosaurs" by Anusuya Chinsamy and Willem J. Hillenius.
- ↑ Pontzer, Herman; Allen, Vivian; Hutchinson, John R. (November 11, 2009). Farke, Andrew Allen (уред.). „Biomechanics of running indicates endothermy in bipedal dinosaurs“. PLOS ONE. San Francisco, CA: PLOS. 4 (11): e7783. Bibcode:2009PLoSO...4.7783P. doi:10.1371/journal.pone.0007783. ISSN 1932-6203. PMC 2772121. PMID 19911059.
- ↑ Smith, Dave; и др. „Hot-Blooded or Cold-Blooded??“. Berkeley: University of California Museum of Paleontology. Посетено на 2019-10-30.
- ↑ Bakker, Robert T. (Spring 1968). Remington, Jeanne E. (уред.). „The Superiority of Dinosaurs“. Discovery: Magazine of the Peabody Museum of Natural History. New Haven, CT: Peabody Museum of Natural History. 3 (2): 11–22. ISSN 0012-3625. OCLC 297237777.
- ↑ 121,0 121,1 Parsons 2001, стр. 22–48, "The Heresies of Dr. Bakker".
- ↑ Perkins, Sid (October 13, 2015). „How to take a dinosaur's temperature“. Science Magazine. Washington, D.C.: American Association for the Advancement of Science. doi:10.1126/science.aad4705. Архивирано од изворникот 2019-09-25. Посетено на 2019-10-30.
- ↑ Farlow & Brett-Surman 1997, стр. 449–473, chpt. 32: "Dinosaurian Physiology: The Case for 'Intermediate' Dinosaurs" by R.E.H. Reid.
- ↑ Ehrlich, Paul R.; Dobkin, David S.; Wheye, Darryl (1988). „Drinking“. Birds of Stanford. Stanford, CA: Stanford University. Посетено на 2007-12-13.
- ↑ Tsahar, Ella; Martínez del Rio, Carlos; Izhaki, Ido; и др. (March 2005). „Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores“ (PDF). The Journal of Experimental Biology. Cambridge: The Company of Biologists. 208 (Pt. 6): 1025–1034. doi:10.1242/jeb.01495. ISSN 0022-0949. PMID 15767304. S2CID 18540594. Архивирано (PDF) од изворникот 2019-10-17. Посетено на 2019-10-31.
- ↑ Skadhauge, Erik; Erlwanger, Kennedy H.; Ruziwa, S.D.; и др. (April 2003). „Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?“. Comparative Biochemistry and Physiology Part A. Amsterdam: Elsevier. 134 (4): 749–755. doi:10.1016/S1095-6433(03)00006-0. ISSN 1095-6433. PMID 12814783.
- ↑ Preest, Marion R.; Beuchat, Carol A. (April 1997). „Ammonia excretion by hummingbirds“. Nature. London: Nature Research. 386 (6625): 561–562. Bibcode:1997Natur.386..561P. doi:10.1038/386561a0. ISSN 0028-0836. S2CID 4372695.
- ↑ Mora, J.; Martuscelli, J; Ortiz Pineda, J.; и др. (July 1965). „The Regulation of Urea-Biosynthesis Enzymes in Vertebrates“. Biochemical Journal. London: Portland Press. 96 (1): 28–35. doi:10.1042/bj0960028. ISSN 0264-6021. PMC 1206904. PMID 14343146.
- ↑ Packard, Gary C. (November–December 1966). „The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates“. The American Naturalist. American Society of Naturalists. 100 (916): 667–682. doi:10.1086/282459. ISSN 0003-0147. JSTOR 2459303.
- ↑ Huxley, Thomas H. (1868). „On the Animals which are most nearly intermediate between Birds and Reptiles“. The Annals and Magazine of Natural History. London: Taylor & Francis. 4 (2): 66–75. Посетено на October 31, 2019.
- ↑ Heilmann 1926
- ↑ Osborn, Henry Fairfield (1924). „Three new Theropoda, Protoceratops zone, central Mongolia“ (PDF). American Museum Novitates. New York: American Museum of Natural History (144): 1–12. ISSN 0003-0082.
- ↑ Ostrom, John H. (1973). „The ancestry of birds“. Nature. London: Nature Research. 242 (5393): 136. Bibcode:1973NPhS..242..136O. doi:10.1038/242136a0. ISSN 0028-0836. S2CID 29873831.
- ↑ Padian 1986, стр. 1–55, "Saurischian Monophyly and the Origin of Birds" by Jacques Gauthier.
- ↑ Mayr, Gerald; Pohl, Burkhard; Peters, D. Stefan (2005). „A Well-Preserved Archaeopteryx Specimen with Theropod Features“. Science. Washington, D.C.: American Association for the Advancement of Science. 310 (5753): 1483–1486. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. ISSN 0036-8075. PMID 16322455. S2CID 28611454.
- ↑ Martin, Larry D. (2006). „A basal archosaurian origin for birds“. Acta Zoologica Sinica. 50 (6): 977–990. ISSN 1674-5507.
- ↑ 137,0 137,1 Feduccia, Alan (October 1, 2002). „Birds are Dinosaurs: Simple Answer to a Complex Problem“. The Auk. Washington, D.C.: American Ornithologists' Union. 119 (4): 1187–1201. doi:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2. ISSN 0004-8038. JSTOR 4090252. Посетено на November 3, 2019.
- ↑ 138,0 138,1 Switek, Brian (July 2, 2012). „Rise of the fuzzy dinosaurs“. News. Nature. London: Nature Research. doi:10.1038/nature.2012.10933. ISSN 0028-0836. S2CID 123219913. Посетено на 2019-01-01.
- ↑ Godefroit, Pascal; Sinitsa, Sofia; Dhouailly, Danielle; Bolotsky, Yuri; Sizov, Alexander (November 2, 2013). Feather-like structures and scales in a Jurassic neornithischian dinosaur from Siberia (PDF). Program and Abstracts of the 73rd Meeting of the Society of Vertebrate Paleontology. Journal of Vertebrate Paleontology. Los Angeles, CA. стр. 135. ISSN 1937-2809. Архивирано од изворникот 2019-02-06. Посетено на 2019-11-01. Supplement to the online Journal of Vertebrate Paleontology, October 2013.
- ↑ Xu, Xing; Norell, Mark A.; Kuang, Xuewen; и др. (2004). „Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids“. Nature. London: Nature Research. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. doi:10.1038/nature02855. ISSN 0028-0836. PMID 15470426. S2CID 4381777.
- ↑ Göhlich, Ursula B.; Chiappe, Luis M. (2006). „A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago“ (PDF). Nature. London: Nature Research. 440 (7082): 329–332. Bibcode:2006Natur.440..329G. doi:10.1038/nature04579. ISSN 0028-0836. PMID 16541071. S2CID 4427002. Архивирано од изворникот (PDF) на April 26, 2019. Посетено на November 1, 2019.
- ↑ Kellner, Alexander W. A.; Wang, Xiaolin; Tischlinger, Helmut; и др. (January 22, 2010). „The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane“. Proceedings of the Royal Society B. London: Royal Society. 277 (1679): 321–329. doi:10.1098/rspb.2009.0846. ISSN 0962-8452. PMC 2842671. PMID 19656798.
- ↑ Wellnhofer, Peter (1988). „A New Specimen of Archaeopteryx“. Science. Washington, D.C.: American Association for the Advancement of Science. 240 (4860): 1790–1792. Bibcode:1988Sci...240.1790W. doi:10.1126/science.240.4860.1790. ISSN 0036-8075. JSTOR 1701652. PMID 17842432. S2CID 32015255.
- —— (1988). „Ein neuer Exemplar von Archaeopteryx“. Archaeopteryx. 6: 1–30.
- ↑ Dong 1992
- ↑ 145,0 145,1 Schweitzer, Mary H.; Watt, J.A.; Avci, R.; и др. (August 15, 1999). „Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid, Shuvuuia deserti“. Journal of Experimental Zoology Part B. Hoboken, NJ: Wiley-Blackwell. 285 (2): 146–157. doi:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A. ISSN 1552-5007. PMID 10440726.
- ↑ Lingham-Soliar, Theagarten (December 2003). „The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers“. Naturwissenschaften. Berlin: Springer Science+Business Media. 90 (12): 563–567. Bibcode:2003NW.....90..563L. doi:10.1007/s00114-003-0483-7. ISSN 0028-1042. PMID 14676953. S2CID 43677545.
- ↑ 147,0 147,1 Feduccia, Alan; Lingham-Soliar, Theagarten; Hinchliffe, J. Richard (November 2005). „Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence“. Journal of Morphology. Hoboken, NJ: John Wiley & Sons. 266 (2): 125–166. doi:10.1002/jmor.10382. ISSN 0362-2525. PMID 16217748. S2CID 15079072.
- ↑ Lingham-Soliar, Theagarten; Feduccia, Alan; Wang, Xiaolin (2007). „A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres“. Proceedings of the Royal Society B. London: Royal Society. 274 (1620): 1823–1829. doi:10.1098/rspb.2007.0352. ISSN 0962-8452. PMC 2270928. PMID 17521978.
- ↑ Prum, Richard O. (2003). „Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002“. The Auk. Washington, D.C.: American Ornithologists' Union. 120 (2): 550–561. doi:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2. ISSN 0004-8038. JSTOR 4090212.
- ↑ Romey, Kristin (December 8, 2016). „First Dinosaur Tail Found Preserved in Amber“. News. Washington, D.C.: National Geographic Society. Архивирано од изворникот 2019-09-24. Посетено на 2019-11-02.
- ↑ Xing, Lida; McKellar, Ryan C.; Xu, Xing; и др. (December 19, 2016). „A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber“. Current Biology. Cambridge, MA: Cell Press. 26 (24): 3352–3360. doi:10.1016/j.cub.2016.10.008. hdl:1983/d3a169c7-b776-4be5-96af-6053c23fa52b. ISSN 0960-9822. PMID 27939315. S2CID 31580099.
- ↑ „Archaeopteryx: An Early Bird“. Berkeley: University of California Museum of Paleontology. Посетено на 2019-10-30.
- ↑ O'Connor, Patrick M.; Claessens, Leon P. A. M. (2005). „Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs“. Nature. London: Nature Research. 436 (7048): 253–256. Bibcode:2005Natur.436..253O. doi:10.1038/nature03716. ISSN 0028-0836. PMID 16015329. S2CID 4390587.
- ↑ Gibson, Andrea (July 13, 2005). „Study: Predatory Dinosaurs had Bird-Like Pulmonary System“. Research Communications. Athens, OH: Ohio University. Посетено на 2019-11-18.[мртва врска]
- ↑ Sereno, Paul C.; Martinez, Ricardo N.; Wilson, Jeffrey A.; и др. (September 2008). Kemp, Tom (уред.). „Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina“. PLOS ONE. San Francisco, CA: PLOS. 3 (9): e3303. Bibcode:2008PLoSO...3.3303S. doi:10.1371/journal.pone.0003303. ISSN 1932-6203. PMC 2553519. PMID 18825273.
- ↑ „Meat-eating dinosaur from Argentina had bird-like breathing system“. University of Michigan News. Ann Arbor, MI: Office of the Vice President for Communications; Regents of the University of Michigan. October 2, 2008. Посетено на November 2, 2019.
- ↑ Xu, Xing; Norell, Mark A. (2004). „A new troodontid dinosaur from China with avian-like sleeping posture“. Nature. London: Nature Research. 431 (7010): 838–841. Bibcode:2004Natur.431..838X. doi:10.1038/nature02898. ISSN 0028-0836. PMID 15483610. S2CID 4362745.
- ↑ Norell, Mark A.; Clark, James M.; Chiappe, Luis M.; и др. (December 28, 1995). „A nesting dinosaur“. Nature. London: Nature Research. 378 (6559): 774–776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0. ISSN 0028-0836. S2CID 4245228.
- ↑ Varricchio, David J.; Moore, Jason R.; Erickson, Gregory M.; и др. (December 19, 2008). „Avian Paternal Care Had Dinosaur Origin“. Science. Washington, D.C.: American Association for the Advancement of Science. 322 (5909): 1826–1828. Bibcode:2008Sci...322.1826V. doi:10.1126/science.1163245. ISSN 0036-8075. PMID 19095938. S2CID 8718747.
- ↑ Wings, Oliver (2007). „A review of gastrolith function with implications for fossil vertebrates and a revised classification“ (PDF). Palaeontologica Polonica. Warsaw: Institute of Paleobiology, Polish Academy of Sciences. 52 (1): 1–16. ISSN 0567-7920. Посетено на November 2, 2019.
- ↑ Dingus & Rowe 1998
- ↑ Le Loeuff, Jean (December 2012). „Paleobiogeography and biodiversity of Late Maastrichtian dinosaurs: how many dinosaur species went extinct at the Cretaceous-Tertiary boundary?“. Bulletin de la Société Géologique de France. Les Ulis: EDP Sciences. 183 (6): 547–559. doi:10.2113/gssgfbull.183.6.547. ISSN 0037-9409.
- ↑ MacLeod, N.; Rawson, P.F.; Forey, P.L.; и др. (1997). „The Cretaceous–Tertiary biotic transition“. Journal of the Geological Society. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. doi:10.1144/gsjgs.154.2.0265. ISSN 0016-7649. S2CID 129654916.
- ↑ Longrich, Nicholas R.; Bhullar, Bhart-Anjan S.; Gauthier, Jacques A. (December 26, 2012). „Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary“. Proc. Natl. Acad. Sci. U.S.A. Washington, D.C.: National Academy of Sciences. 109 (52): 21396–21401. Bibcode:2012PNAS..10921396L. doi:10.1073/pnas.1211526110. ISSN 0027-8424. PMC 3535637. PMID 23236177.
- ↑ Keller, Gerta; Adatte, Thierry; Gardin, Silvia; и др. (April 30, 2008). „Main Deccan volcanism phase ends near the K–T boundary: Evidence from the Krishna–Godavari Basin, SE India“. Earth and Planetary Science Letters. Amsterdam: Elsevier. 268 (3–4): 293–311. Bibcode:2008E&PSL.268..293K. doi:10.1016/j.epsl.2008.01.015. ISSN 0012-821X.
- ↑ Mullen, Leslie (October 20, 2004). „Multiple Impacts?“. Astrobiology Magazine. Washington, D.C.: NASA. ISSN 2152-1239. Посетено на 2019-11-03.
- ↑ Randall 2015
- ↑ Alvarez, L.W.; Alvarez, W.; Asaro, F.; Michel, H.V. (1980). „Extraterrestrial Cause for the Cretaceous-Tertiary Extinction“ (PDF). Science. 208 (4448): 1095–1108. Bibcode:1980Sci...208.1095A. CiteSeerX 10.1.1.126.8496. doi:10.1126/science.208.4448.1095. ISSN 0036-8075. PMID 17783054. S2CID 16017767. Архивирано од изворникот (PDF) на July 8, 2010. Посетено на October 30, 2019.
- ↑ Pope, K.O.; Ocampo, A.C.; Kinsland, G.L.; и др. (1996). „Surface expression of the Chicxulub crater“. Geology. Boulder, CO: Geological Society of America. 24 (6): 527–530. Bibcode:1996Geo....24..527P. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. ISSN 0091-7613. PMID 11539331.
- ↑ Robertson, Douglas S.; McKenna, Malcolm C.; Toon, Owen B.; и др. (May–June 2004). „Survival in the first hours of the Cenozoic“ (PDF). Geological Society of America Bulletin. Boulder, CO: Geological Society of America. 116 (5–6): 760–768. Bibcode:2004GSAB..116..760R. doi:10.1130/B25402.1. ISSN 0016-7606. Архивирано од изворникот (PDF) на 2012-09-18. Посетено на 2011-06-15.
- ↑ Hotz, Robert Lee (September 15, 2019). „Details Discovered About the Day the Dinosaurs Died“. News. MSN.com. Redmond, WA: Microsoft. Архивирано од изворникот 2019-09-18. Посетено на 2019-11-08.
- —— (September 9, 2019). „Scientists Discover New Evidence of the Asteroid That Killed Off the Dinosaurs“. The Wall Street Journal. New York: Dow Jones & Company. ISSN 0099-9660. Архивирано од изворникот 2019-11-08. Посетено на 2019-11-08.
- ↑ "What Really Killed the Dinosaurs?". Horizon. BBC Television. No. 4.
- ↑ Alvarez 1997, стр. 130–146, chpt. 7: "The World after Chicxulub".
- ↑ Fassett, J.E.; Heaman, L.M.; Simonetti, A. (2011). „Direct U–Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico“. Geology. 39 (2): 159–162. Bibcode:2011Geo....39..159F. doi:10.1130/G31466.1. ISSN 0091-7613.
- ↑ Fassett, J.E.; Heaman, L.M.; Simonetti, A. (2009). „New geochronologic and stratigraphic evidence confirms the Paleocene age of the dinosaur-bearing Ojo Alamo Sandstone and Animas Formation in the San Juan Basin, New Mexico and Colorado“. Palaeontologia Electronica. 12 (1): 3A.
- ↑ Sloan, R.E.; Rigby, J.K. Jr.; Van Valen, L.M.; и др. (1986). „Gradual Dinosaur Extinction and Simultaneous Ungulate Radiation in the Hell Creek Formation“. Science. 232 (4750): 629–633. Bibcode:1986Sci...232..629S. doi:10.1126/science.232.4750.629. ISSN 0036-8075. PMID 17781415. S2CID 31638639.
- ↑ Kubo, Tai; Benton, Michael J. (November 2007). „Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates“. Palaeontology. Hoboken, NJ: Wiley-Blackwell. 50 (6): 1519–1529. doi:10.1111/j.1475-4983.2007.00723.x. ISSN 0031-0239.
- ↑ „Dinosaur bones 'used as medicine'“. BBC News. London: BBC. July 6, 2007. Архивирано од изворникот August 27, 2019. Посетено на November 4, 2019.
- ↑ Paul 2000, стр. 10–44, chpt. 1: "A Brief History of Dinosaur Paleontology" by Michael J. Benton.
- ↑ 180,0 180,1 Farlow & Brett-Surman 1997, стр. 3–11, chpt. 1: "The Earliest Discoveries" by William A.S. Sarjeant.
- ↑ Plot 1677
- ↑ Plot 1677, стр. 136
- ↑ „Robert Plot“ (PDF). Learning more. Oxford: Oxford University Museum of Natural History. 2006. Архивирано од изворникот (PDF) на 2006-10-01. Посетено на 2019-11-14.
- ↑ Lhuyd 1699, стр. 67
- ↑ Delair, Justin B.; Sarjeant, William A.S. (2002). „The earliest discoveries of dinosaurs: the records re-examined“. Proceedings of the Geologists' Association. Amsterdam: Elsevier on behalf of the Geologists' Association. 113 (3): 185–197. doi:10.1016/S0016-7878(02)80022-0. ISSN 0016-7878.
- ↑ Gunther 1968
- ↑ Buckland, William (1824). „Notice on the Megalosaurus or great Fossil Lizard of Stonesfield“. Transactions of the Geological Society of London. London: Geological Society of London. 1 (2): 390–396. doi:10.1144/transgslb.1.2.390. ISSN 2042-5295. S2CID 129920045. Архивирано (PDF) од изворникот October 21, 2019. Посетено на November 5, 2019.
- ↑ Mantell, Gideon A. (1825). „Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex“. Philosophical Transactions of the Royal Society of London. London: Royal Society. 115: 179–186. Bibcode:1825RSPT..115..179M. doi:10.1098/rstl.1825.0010. ISSN 0261-0523. JSTOR 107739.
- ↑ Farlow & Brett-Surman 1997, стр. 14, chpt. 2: "European Dinosaur Hunters" by Hans-Dieter Sues.
- ↑ Rupke 1994
- ↑ Prieto-Marquez, Albert; Weishampel, David B.; Horner, John R. (March 2006). „The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus“ (PDF). Acta Palaeontologica Polonica. Warsaw: Institute of Paleobiology, Polish Academy of Sciences. 51 (1): 77–98. ISSN 0567-7920. Архивирано (PDF) од изворникот 2019-06-22. Посетено на 2019-11-05.
- ↑ Holmes 1998
- ↑ Salgado, Leonardo; Gasparini, Zulma (March 2006). „Reappraisal of an ankylosaurian dinosaur from the Upper Cretaceous of James Ross Island (Antarctica)“ (PDF). Geodiversitas. Paris: Muséum national d’Histoire naturelle. 28 (1): 119–135. ISSN 1280-9659. Архивирано (PDF) од изворникот 2018-05-02. Посетено на 2019-11-05.
- ↑ 194,0 194,1 Bakker 1986
- ↑ Bakker, R.T. (1968). „The Superiority of Dinosaurs“. Discovery: Magazine of the Peabody Museum of Natural History. 3 (2): 11–22. ISSN 0012-3625. OCLC 297237777.
- ↑ Bakker, R.T. (1972). „Anatomical and Ecological Evidence of Endothermy in Dinosaurs“. Nature. 238 (5359): 81–85. doi:10.1038/238081a0. S2CID 4176132.
- ↑ 197,0 197,1 Taylor, M.P. (2010). „Sauropod dinosaur research: a historical review“. Geological Society, London, Special Publications. 343 (1): 361–386. doi:10.1144/SP343.22. S2CID 910635.
- ↑ Arbour, Victoria (2018). „Results roll in from the dinosaur renaissance“. Science. 360 (6389): 611. doi:10.1126/science.aat0451. S2CID 46887409.
- ↑ Naish, D. (2009). The Great Dinosaur Discoveries. London, UK: A & C Black Publishers Ltd. стр. 89–93. ISBN 978-1-4081-1906-8.
- ↑ Evershed, Nick (May 1, 2009). „Blood, tissue extracted from duck-billed dinosaur bone“. Cosmos Online. Adelaide: Cosmos Media Pty Ltd. ISSN 1832-522X. Архивирано од изворникот на 2016-02-15. Посетено на 2013-10-02.
- ↑ Schweitzer, Mary H.; Wenxia, Zheng; Cleland, Timothy P.; и др. (January 2013). „Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules“. Bone. Amsterdam: Elsevier. 52 (1): 414–423. doi:10.1016/j.bone.2012.10.010. ISSN 8756-3282. PMID 23085295.
- ↑ Wang, Hai-Lin; Yan, Zi-Ying; Jin, Dong-Yan (1997). „Reanalysis of Published DNA Sequence Amplified from Cretaceous Dinosaur Egg Fossil“ (PDF). Molecular Biology and Evolution. Oxford: Oxford University Press on behalf of the Society for Molecular Biology and Evolution. 14 (5): 589–591. doi:10.1093/oxfordjournals.molbev.a025796. ISSN 0737-4038. PMID 9159936. Архивирано (PDF) од изворникот July 30, 2018. Посетено на November 7, 2019.
- ↑ Chang, Belinda S. W.; Jönsson, Karolina; Kazmi, Manija A.; и др. (2002). „Recreating a Functional Ancestral Archosaur Visual Pigment“. Molecular Biology and Evolution. Oxford: Oxford University Press on behalf of the Society for Molecular Biology and Evolution. 19 (9): 1483–1489. doi:10.1093/oxfordjournals.molbev.a004211. ISSN 0737-4038. PMID 12200476.
- ↑ Schweitzer, Mary H.; Marshall, Mark; Carron, Keith; и др. (1997). „Heme compounds in dinosaur trabecular bone“. Proc. Natl. Acad. Sci. U.S.A. Washington, D.C.: National Academy of Sciences. 94 (12): 6291–6296. Bibcode:1997PNAS...94.6291S. doi:10.1073/pnas.94.12.6291. ISSN 0027-8424. PMC 21042. PMID 9177210.
- ↑ Embery, Graham; Milner, Angela C.; Waddington, Rachel J.; и др. (2003). „Identification of Proteinaceous Material in the Bone of the Dinosaur Iguanodon“. Connective Tissue Research. Milton Park, Oxfordshire: Taylor & Francis. 44 (1): 41–46. doi:10.1080/03008200390152070. ISSN 1607-8438. PMID 12952172. S2CID 2249126.
- ↑ Peterson, Joseph E.; Lenczewski, Melissa E.; Scherer, Reed P. (October 12, 2010). Stepanova, Anna (уред.). „Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs“. PLOS ONE. San Francisco, CA: PLOS. 5 (10): e13334. Bibcode:2010PLoSO...513334P. doi:10.1371/journal.pone.0013334. ISSN 1932-6203. PMC 2953520. PMID 20967227.
- ↑ Bertazzo, Sergio; Maidment, Susannah C.R.; Kallepitis, Charalambos; и др. (June 9, 2015). „Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens“. Nature Communications. London: Nature Research. 6: 7352. Bibcode:2015NatCo...6.7352B. doi:10.1038/ncomms8352. ISSN 2041-1723. PMC 4468865. PMID 26056764.
- ↑ Mortillaro, Nicole (June 9, 2015). „Scientists discover 75-million-year-old dinosaur blood and tissue“. Global News. Toronto: Corus Entertainment. Архивирано од изворникот 2019-06-11. Посетено на 2019-11-07.
- ↑ Предлошка:Cite Merriam-Webster
- ↑ Sarjeant 1995, стр. 255–284, chpt. 15: "The Dinosaurs and Dinomania over 150 Years" by Hugh S. Torrens.
- ↑ Currie & Padian 1997, стр. 347–350, "History of Dinosaur Discoveries: First Golden Period" by Brent H. Breithaupt.
- ↑ Dickens 1853, стр. 1, chpt. I: "London. Michaelmas Term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborn Hill."
- ↑ Farlow & Brett-Surman 1997, стр. 675–697, chpt. 43: "Dinosaurs and the Media" by Donald F. Glut and M.K. Brett-Surman.
- ↑ Italo Kalvino, Kosmikomike: stare i nove. Beograd: Paidea, 2008, стр. 75-88.
- ↑ Хигсов Бозон, Педерски Катахрезис, Темплум, Скопје, 2011.
- ↑ "Walk the Dinosaur" Was (Not Was) (пристапено на 8.11.2017)
- ↑ IMDb, The Good Dinosaur (2015) (пристапено на 8.3.2017)
Библиографија
уреди- Alvarez, Walter (1997). T. rex and the Crater of Doom. Princeton, NJ: Princeton University Press. ISBN 978-0-691-01630-6. LCCN 96049208. OCLC 1007846558. Посетено на November 4, 2019.
- Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction. New York: William Morrow and Company. ISBN 978-0-688-04287-5. LCCN 86012643. OCLC 13699558. Посетено на November 6, 2019.
- Benton, Michael J. (2005). Vertebrate Palaeontology (3. изд.). Malden, MA: Blackwell Publishing. ISBN 978-0-632-05637-8. LCCN 2003028152. OCLC 53970617. Посетено на October 30, 2019.
- Brusatte, Stephen L. (2012). Benton, Michael J. (уред.). Dinosaur Paleobiology. Topics in Paleobiology. Foreword by Michael J. Benton. Hoboken, NJ: Wiley-Blackwell. ISBN 978-0-470-65658-7. LCCN 2011050466. OCLC 781864955. Посетено на October 30, 2019.
- Chiappe, Luis M.; Witmer, Lawrence M., уред. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. ISBN 978-0-520-20094-4. LCCN 2001044600. OCLC 901747962.
- Colbert, Edwin H. (1971) [Originally published, New York: E. P. Dutton, 1968; London: Evans Brothers Ltd, 1969]. Men and Dinosaurs: The Search in Field and Laboratory. Harmondsworth: Penguin. ISBN 978-0-14-021288-4. OCLC 16208760. Посетено на October 31, 2019.
- Cowen, Richard (2005). History of Life (4. изд.). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-1756-2. LCCN 2003027993. OCLC 53970577. The 5th edition of the book is available from the Internet Archive. Retrieved 2019-10-19.
- Currie, Philip J.; Padian, Kevin, уред. (1997). Encyclopedia of Dinosaurs. San Diego, CA: Academic Press. ISBN 978-0-12-226810-6. LCCN 97023430. OCLC 436848919. Посетено на October 30, 2019.
- Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; Wright, Joanna L., уред. (2004). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Life of the Past. Bloomington, IN: Indiana University Press. ISBN 978-0-253-34373-4. LCCN 2003019035. OCLC 52942941.
- Curry Rogers, Kristina A.; Wilson, Jeffrey A., уред. (2005). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. ISBN 978-0-520-24623-2. LCCN 2005010624. OCLC 879179542.
- Desmond, Adrian J. (1975). The Hot-Blooded Dinosaurs: A Revolution in Palaeontology. London: Blond & Briggs. ISBN 978-0-8037-3755-6. LCCN 76359907. OL 4933052M. Посетено на October 30, 2019.
- Dickens, Charles (1853). Bleak House. London: Bradbury and Evans. Посетено на November 7, 2019.
- Dodson, Peter; Gingerich, Philip D., уред. (1993). „Functional Morphology and Evolution“. The American Journal of Science and Arts. A special volume of the American Journal of Science. New Haven, CT: Kline Geology Laboratory, Yale University. 293-A. ISSN 0002-9599. OCLC 27781160.
- Dong, Zhiming (1992). Dinosaurian Faunas of China (English. изд.). Beijing; Berlin; New York: China Ocean Press; Springer-Verlag. ISBN 978-3-540-52084-9. LCCN 92207835. OCLC 26522845.
- Dyke, Gareth; Kaiser, Gary, уред. (2011). Living Dinosaurs: The Evolutionary History of Modern Birds. Chichester; Hoboken, NJ: Wiley-Blackwell. ISBN 978-0-470-65666-2. LCCN 2010043277. OCLC 729724640.
- Farlow, James O.; Brett-Surman, M.K., уред. (1997). The Complete Dinosaur. Bloomington, IN: Indiana University Press. ISBN 978-0-253-33349-0. LCCN 97-23698. OCLC 924985811. Посетено на October 14, 2019.
- Foster, John R.; Lucas, Spencer G., уред. (2006). „Paleontology and Geology of the Upper Jurassic Morrison Formation“. Bulletin of the New Mexico Museum of Natural History and Science. New Mexico Museum of Natural History and Science Bulletin. Albuquerque, NM: New Mexico Museum of Natural History and Science. 36. ISSN 1524-4156. OCLC 77520577. Посетено на October 21, 2019.
- Glut, Donald F. (1997). Dinosaurs: The Encyclopedia. Foreword by Michael K. Brett-Surman. Jefferson, NC: McFarland & Company. ISBN 978-0-89950-917-4. LCCN 95047668. OCLC 33665881.
- Gunther, Robert Theodore, уред. (1968) [First printed in Oxford 1945]. Life and Letters of Edward Lhwyd. Early Science in Oxford. XIV. Preface by Albert Everard Gunther (Reprint. изд.). London: Dawsons of Pall Mall. ISBN 978-0-7129-0292-2. LCCN 22005926. OCLC 43529321. Посетено на November 4, 2019.
- Hansell, Mike (2000). Bird Nests and Construction Behaviour. Pen and ink illustration by Raith Overhill. Cambridge: University of Cambridge Press. ISBN 978-0-521-46038-5. LCCN 99087681. OCLC 876286627. Посетено на October 30, 2019.
- Heilmann, Gerhard (1926). The Origin of Birds. London; New York: H. F. & G. Witherby; D. Appleton & Company. LCCN 27001127. OCLC 606021642.
- Holmes, Thom (1998). Fossil Feud: The Rivalry of the First American Dinosaur Hunters. Parsippany, NJ: Julian Messner. ISBN 978-0-382-39149-1. LCCN 96013610. OCLC 34472600.
- Holtz, Thomas R. Jr. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Illustrated by Luis V. Rey. New York: Random House. ISBN 978-0-375-82419-7. LCCN 2006102491. OCLC 77486015. Посетено на October 22, 2019.
- Lambert, David; The Diagram Group (1990). The Dinosaur Data Book: The Definitive, Fully Illustrated Encyclopedia of Dinosaurs. New York: Avon Books. ISBN 978-0-380-75896-8. LCCN 89092487. OCLC 21833417. Посетено на October 14, 2019.
- Lessem, Don; Glut, Donald F. (1993). The Dinosaur Society's Dinosaur Encyclopedia. Illustrations by Tracy Lee Ford; scientific advisors, Peter Dodson, et al. New York: Random House. ISBN 978-0-679-41770-5. LCCN 94117716. OCLC 30361459. Посетено на October 30, 2019.
- Lhuyd, Edward (1699). Lithophylacii Britannici ichnographia [British figured stones]. London: Ex Officina M.C. Архивирано од изворникот на 2020-08-14. Посетено на November 4, 2019.
- Mayr, Gerald (2009). Paleogene Fossil Birds. Berlin: Springer-Verlag. doi:10.1007/978-3-540-89628-9. ISBN 978-3-540-89627-2. LCCN 2008940962. OCLC 916182693. Посетено на October 30, 2019.
- Norell, Mark; Gaffney, Eugene S.; Dingus, Lowell (2000) [Originally published as Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf, 1995]. Discovering Dinosaurs: Evolution, Extinction, and the Lessons of Prehistory (Revised. изд.). Berkeley: University of California Press. ISBN 978-0-520-22501-5. LCCN 99053335. OCLC 977125867. Посетено на October 30, 2019.
- Olshevsky, George (2000). An Annotated Checklist of Dinosaur Species by Continent. Mesozoic Meanderings. 3. Illustrated by Tracy Lee Ford. San Diego, CA: Publications Requiring Research. ISSN 0271-9428. LCCN 00708700. OCLC 44433611.
- Owen, Richard (1842). „Report on British Fossil Reptiles. Part II“. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841. London: John Murray. стр. 60–204. ISBN 978-0-8201-1526-9. LCCN 99030427. OCLC 1015526268. Посетено на October 13, 2019.
- Padian, Kevin, уред. (1986). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences. 8. San Francisco, CA: California Academy of Sciences. ISBN 978-0-940228-14-6. OCLC 946083441. OL 9826926M.
- Parsons, Keith M. (2001). Drawing out Leviathan: Dinosaurs and the Science Wars. Life in the Past. Bloomington, IN: Indiana University Press. ISBN 978-0-253-33937-9. LCCN 2001016803. OCLC 50174737. Посетено на October 30, 2019.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World: A Complete Illustrated Guide. New York: Simon & Schuster. ISBN 978-0-671-61946-6. LCCN 88023052. OCLC 859819093. Посетено на October 30, 2019.
- Paul, Gregory S., уред. (2000). The Scientific American Book of Dinosaurs (1. изд.). New York: St. Martin's Press. ISBN 978-0-312-26226-6. LCCN 2001269051. OCLC 45256074.
- Paul, Gregory S. (2010). The Princeton Field Guide to Dinosaurs. Princeton Field Guides. Princeton, NJ: Princeton University Press. ISBN 978-0-691-13720-9. LCCN 2010014916. OCLC 907619291.
- Plot, Robert (1677). The Natural History of Oxford-shire: Being an Essay toward the Natural History of England. Printed at the Theater in OXFORD, and are to be had there: And in London at Mr. S. Millers, at the Star near the West-end of St. Pauls Church-yard. Oxford; London. LCCN 11004267. OCLC 933062622. Посетено на November 13, 2019.
- Randall, Lisa (2015). Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the Universe. New York: HarperCollins: Ecco. ISBN 978-0-06-232847-2. LCCN 2016427646. OCLC 962371431.
- Rupke, Nicolaas A. (1994). Richard Owen: Victorian Naturalist. New Haven: Yale University Press. ISBN 978-0-300-05820-8. LCCN 93005739. OCLC 844183804. Посетено на November 5, 2019.
- Sarjeant, William A.S., уред. (1995). Vertebrate Fossils and the Evolution of Scientific Concepts: Writings in Tribute to Beverly Halstead, by Some of His Many Friends. Modern Geology. Amsterdam: Gordon and Breach Publishers. ISBN 978-2-88124-996-9. ISSN 0026-7775. LCCN 00500382. OCLC 34672546. "Reprint of papers published in a special volume of Modern geology [v. 18 (Halstead memorial volume), 1993], with five additional contributions.--Pref."
- Tanner, Lawrence H.; Spielmann, Justin A.; Lucas, Spencer G., уред. (2013). „The Triassic System: New Developments in Stratigraphy and Paleontology“. Bulletin of the New Mexico Museum of Natural History and Science. New Mexico Museum of Natural History and Science Bulletin. Albuquerque, NM: New Mexico Museum of Natural History and Science. 61. ISSN 1524-4156. OCLC 852432407. Посетено на October 21, 2019.
- Weishampel, David B.; Dodson, Peter; Osmólska, Halszka, уред. (2004). The Dinosauria (2. изд.). Berkeley: University of California Press. ISBN 978-0-520-25408-4. LCCN 2004049804. OCLC 154697781.
Надворешни врски
уреди| „Диносаурус“ на Ризницата ? |
| „Dinosauria“ на Викивидовите ? |
Генерално
- „DinoDatabase.com“ – Стотици диносауруси и теми поврзани со диносауруси.
Слики
- „gspauldino.com“ – Анатомија и слики на влијателен палеонтолог.
- „Scott Hartman's Skeletal Drawing.com“ – Професионални реставрации на бројни диносауруси и дискусии за анатомијата на диносаурусите.
- „Dinosaur Discovery - Early Published Images“ Архивирано на 15 август 2020 г. – Колекција на слики од раните дела на диносаурусите во Библиотеката на Линда Хол, како поддршка на изложбата, Paper Dinosaurs, 1824–1969 Архивирано на 15 август 2020 г..
Видео
- BBC Nature: Prehistoric Life: Dinosaurs – Реконструкции и стручни толкувања, вклучително и Одењето со диносауруси. (Архивирано мрежно место последен пат подновено од БиБиСи во октомври 2014 г.)
- BBC Explainer: Dinosaurs – Анимација Комплетна историја во 4 минути.
- "The origin, evolution, and extinction of the dinosaurs" – Видео од Стивен Брусати, 15 април, 2014.
Популарно
- „The Dino Directory“ – Добро илустриран директориум за диносауруси од Музејот на природна историја во Лондон.
- Dinosaurnews Архивирано на 25 јануари 2021 г. – Наслови поврзани со диносаурусите од целиот свет, вклучително и откритија и многу врски.
- „The Dinosauria“ – Од Музејот на палеонтологија на Универзитетот во Калифорнија.
- „Zoom Dinosaurs“ – Од страницата Enchanted Learning. Kids, информативни страници и статистики, теории, историја.
Технички
- Palaeontologia Electronica. Coquina Press. Технички дневник на Интернет.