Мутациски потписи
Мутациски потписи ― карактеристични комбинации на видови мутации кои произлегуваат од специфични постапки на мутагенеза како што се забегување на репликацијата на ДНК, егзогени и ендогени изложувања на генотоксин, неисправни патишта на поправка на ДНК и ензимско уредување на ДНК.[1]
Поимот е користен за два различни концепти, често комбинирани: потписи на мутагени и туморски потписи. Неговата првична употреба, потпис на мутаген, се однесува на шема на мутации направени во лабораторија од познат мутаген, а не направена од други мутагени – единствена за мутагенот како што човечкиот потпис е единствен за потписникот. Уникатноста овозможува мутагенот да биде изведен од мутациите на клетката.[2] Подоцна, фразата се однесувала на модел на мутации карактеристични за видот тумор, иако обично не е единствен за видот тумор ниту за мутаген.[3][4] Ако мутациониот потпис на туморот се совпаѓа со единствениот мутагенски мутациски потпис, валидно е да биде заклучена изложеноста на канцерогенот или постапката на мутагенеза што била настанато во далечното минато на пациентот.[2] Сè повеќе рафинираните туморски потписи стануваат доделени на мутагени потписи.[5]
Дешифрирањето на мутациските потписи кај ракот дава увид во биолошките механизми вклучени во канцерогенезата и нормалната соматска мутагенеза.[6] Мутациските потписи ја покажале својата применливост во лекувањето на ракот и спречувањето рак. Напредокот во областите на онкогеномиката овозможиле развој и употреба на молекуларно насочена терапија, но таквите терапии историски се насочувале на инхибиција на онкогени двигатели (на пр. мутација на добивка на функција на рецептор на епидермален фактор на раст и третман со инхибитор на рецептор на епидермален фактор на раст кај рак на дебелото црево[7]). Во поново време, профилирањето на мутациски потписи било покажано успешно во насочувањето на онколошкото раководење и употребата на целни терапии (на пр. имунотерапија во поправка на неусогласеност на различни видови рак,[8] платина и инхибитор на поли АДП-рибозна полимераза да се искористи синтетичката смртност при недостаток на хомологна рекомбинација во ракот на дојката).[9]
Општи концепти
уреди
Механизми - преглед
уредиБиолошките механизми на мутагенезата подлежат на мутациски потписи (на пр. потписите 1 до 30 на COSMIC) вклучуваат, но не се ограничени на:[б 1][4]
- Забегување во репликацијата на ДНК
- Генотоксини
- Ендегени клеточни мутации (на пример, спонтаната деаминација на 5-метилцитозин води до преод C>T
- Екзогени/ракородни
- Ултравиолетово зрачење: зрачењето предизвикува директна оштетување на ДНК и е познат фактор на ризик за рак на кожата (на пример, меланом)
- Алкилирачки антинеопластични агенси: Оваа група хемиотерапевтични агенси додава алкилна група на ДНК, што предизвикува крстосврзување на ДНК и мешање во репликацијата на ДНК и поправката на ДНК. Клетки на ракот се најзатежени поради нивната висока митоза.
- Тутун: тутунот содржи неколку канцерогени кои се штетни за ДНК, вклучувајќи полициклични ароматски јаглеводороди, акролеин, нитрозамини, цијанид и друго.
- Недостаток на поправка на ДНК
- Недостиг на хомологна рекомбинација: прекинот на ДНК-то со двојно влакно бара хомоложен механизам на рекомбинација за точен исправка на точките на прекин.
- Негостиг на поправка на несогласување на ДНК: Машинеријата за поправка на несогласување препознава и го поправува погрешно вметнување, бришење или погрешно вметнување на базен пар.
- Ензимско уредување на ДНК
- Ензими на цитидинска деаминаза: Ова семејство ензими се дел од вродениот имунолошки систем и се вклучени во контрола на ретровируси и транспозони (вклучувајќи ендогени ретровируси). Овие ензими (цитидинска деаминаза, активирана цитидинска декаманиза и белковинското семејство како што е аполипобелковина Б на ензим на информациска РНК за уредување, каталитички полипептид) активно предизвикуваат деаминација на цитидин и со тоа воведуваат C> T преодни мутации.
Геномски податоци
уредиАнализата на мутациските потписи на ракот бара геномски податоци од секвенционирање на геномот на ракот со спарено-нормално секвенционирање на ДНК со цел да биде создаден каталог на туморските мутации (видови и бројки на мутации) на одреден тумор. Различни видови мутации (на пример, еднократни нуклеотидни варијанти, индели, структурни варијанти) можат да бидат користени поединечно или во комбинација за моделирање на мутациски потписи во ракот.
Видови мутации: базни замени
уредиПостојат шест класи базни замени: C>A, C>G, C>T, T>A, T>C, T>G. Замена на G>T е сметана за еднаква замена на C>A бидејќи не е можно да биде разликувана на која нишка на ДНК (напред или назад) првично била сторена замена. Затоа, и замените C>A и G>T се сметани како дел од класата „C>A“. Поради истата причина, мутациите G>C, G>A, A>T, A>G и A>C се броени како дел од класите „C>G“, „C>T“, „T>A“, „T >C“ и „T>G“ соодветно.
Приемањето на информациите од соседните бази 5' и 3' (исто така наречени крилни базни парови или тринуклеотиден контекст) доведува до 96 можни видови мутации (на пример, A[C>A]A, A[ C>A]T, итн.). Каталогот за мутации на тумор е создаван со категоризација на секоја една нуклеотидна варијанта (синоними: замена на база-пар или замена на точкеста мутација) во еден од 96 видови мутации и пресметување на вкупниот број на заменети за секој од овие 96 видови мутација (види слика).
Каталог на туморски мутации
уреди
Откако ќе биде добиен каталогот за мутации (на пример, бројот на сите 96 видови мутации) на тумор, постојат два пристапи за дешифрирање на придонесите на различни мутациски потписи на туморскиот геномски предел:
- Каталогот на мутации на туморот е споредуван со референтен каталог на мутации, или референтен набор на податоци за мутации, како што се 21 потписи на мутациски постапки во човечкиот рак[4] од базата на податоци Каталог на соматски мутации во ракот (COSMIC).[1]
- Моделирањето отпочеток на мутациските потписи може да биде постигнато со користење на статистички методи како што е не-негативна факторизација на матрицата за да бидат идентификувани потенцијални нови мутациски постапки.[10]
Идентификувањето на придонесите на различни мутациски потписи за канцерогенеза дава увид во туморската биологија и може да понуди можности за целна терапија.
Видови мутации: индели
уредиПотпис 3, забележан во тумор со недостиг на хомологна рекомбинација, е поврзан со зголеменото оптоварување на големи индели (до 50 нуклеотиди) со преклопување на микрохомологијата на точките на прекин.[4] Во такви тумори, прекините на двојните влакна на ДНК се поправани со неточни механизми за поправка на нехомологната крајна поврзаност или микрохомолошка посредна крајна поврзаност наместо високо достапната хомологна рекомбинација.
Потпис 6, забележан во туморите со микросателитна нестабилност, исто така содржи збогатување на 1bp индели во региони на повторување на нуклеотидите.
Видови мутации: структурни варијанти
уредиНедостатокот на хомологната рекомбинација води до модел на замена на потписот 3, но исто така до зголемување на оптоварот на структурните варијанти. Во отсуство на хомологна рекомбинација, нехомологната крајна поврзаност води до големи структурни варијанти како што се хромозомни транслокации, хромозомски инверзии и варијанти на бројот на копии.
Мутациски потписи
уредиНекои потписи се сеприсутни во различни видови рак (на пример, потпис 1) додека некои други имаат тежнеење да се поврзуваат со специфични ракови (на пример , потпис 9 и лимфоидни малигни тумори).[4]
Некои мутациски потписи се одликуваат со силна транскрипцисна пристрасност со замени кои пожелно влијаат на една од нишките на ДНК, или транскрибираната или нетранскрибираната низа (Потписи 5, 7, 8, 10, 12, 16).[4]
Мутагенеза поврзана со возраста
уредиПотписот 1 покажува преовладување на C>T преодот во Np[C> T]G тринуклеотидни контексти и е корелирано со возраста на пациентот во време на дијагнозата на рак. Основниот предложен биолошки механизам е спонтаното деаминирање на 5-метилцитозин.[4]
Потписот 5 има преовладување на T>C замени во контекстот на ApTpN тринуклеотидот со претежност на транскрипциските нишки.[6]
Недостиг на хомологна рекомбинација
уредиПотписот 3 покажува високи бројки на мутации на повеќе класи на мутации и е поврзан со зародишни и соматски мутации на гените BRCA1 и BRCA2 во неколку видови рак (на пример, дојка, панкреас, јајници, простата). Овој знак е резултат на недостаток на поправка на ДНК на преки на двојно влакно (или недостаток на хомологна рекомбинација). Потписот 3 е поврзан со висок товар на инделите со микрохомологија на точките на прекин.[6]
Ензими APOBEC
уредиСемејството APOBEC3 на ензими на цитидинска деаминаза реагира на вирусни инфекции со уредување на вирусниот геном, но ензимската активност на APOBEC3A и APOBEC4B исто така е откриена дека предизвикува несакано уредување на геномот на домаќинот и може дури да учествува во онкогенезата во раковите поврзани со човечкиот папиломен вирус.[11]
Потписите 2 и 13 се збогатени за C>T и C>G замени и се сметани дека произлегуваат од активностите на цитидинската деаминаза на семејството на ензимите со активираната цитидинска деаминаза/APOBEC.[6]
Полиморфизмот на бришење на зародишите, кој вклучува APOBEC3A и APOBEC 3B е поврзан со висока тегоба на мутациите на потписите 2 и 13.[12] Овој полиморфизам е сметан за умерен (двапати повисоки од ризикот од позадина) за ризик од рак на дојка.[13] Точните улоги и механизми кои лежат зад уредување на геномите со посредство на APOBEC се уште не се целосно дефинирани, но се смета дека комплексот активираната цитидинска деаминаза/APOBEC е вклучен во имунолошкиот одговор на домаќинот на вирусни инфекции и метаболизмот на липидите.[14]
Во потписите 2 и 13 има цитозин за замената на урацил поради цитидинските деаминази. Потписот 2 има поголем процент на C[T>C]N заменети и потписот 13 има поголем процент на T[C>G]N замените. Мутагенезата која е посредувана со APOBEC3A и APOBEC3B, претпочитувачки вклучува бавна нишка на ДНК за време на репликацијата.[15]
Недостиг на неусогласена поправка
уредиЧетири мутациски потписи на базата на податоци COSMIC, се поврзани со недостиг на неусогласена поправка на ДНК и се наоѓаат во тумори со микросателитна нестабилност: потпис 6, 15, 20 и 26.[6] Губењето на функцијата на гените MLH1, MSH2, MSH6 или PMS2 предизвикуваат дефектна поправка на ДНК несогласувања.
Поправка на читање на ДНК
уредиПотписот 10 има транскрипцијална пристрасност и е збогатен за C>A заменети во контекстот на TpCpT, како и за T>G замени во контекст на tpTpTp.[6] Потписот 10 е поврзан со променена функција на полимеразата епсилон на ДНК, што резултира со појава на недостаток на поправка на читање на ДНК. И зародишните и соматските мутации на егзонуклеазата на генот POLE се поврзани со потписот 10.[16]
Поправка со вадење на базата
уреди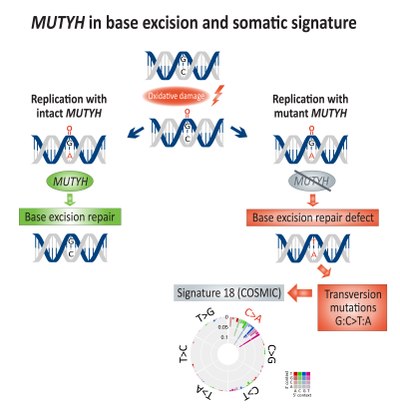
Соматското збогатување за трансверзни мутации (G:C>T:A) е поврзано со недостаток на поправка со вадење на базата и поврзано со дефектниот MUTYH, гликозилаза на ДНК, во ракот на дебелото црево.[17] Директната оштетување на оксидацијата на ДНК води до создавање на 8-оксигуанин, кој ако остане непоправен, ќе доведе до вклучување на аденин наместо цитозин за време на репликацијата на ДНК.[18] MUTYH го кодира ензимот mutY аденин гликозилаза кој го извлекува несогласениот аденоин од 8-оксигуанин:аденоинската база, со што овозможува механизми за поправка на ДНК кои вклучуваат OGG1 (оксигуанин гликозилаза) и NUDT1 (нудикс хидролаза 1, исто така познат како MTH1, MutT хомолог 1) да го отстранат оштетениот 8-оксиуанин.
Изложеност на егзогени генотоксини
уредиИзбраните егзогени генотоксини/канцерогени и нивните мутагенско предизвикани оштетувања и механизми за поправка на ДНК се поврзани со специфични молекуларни потписи.
Ултравиолетовo зрачење
уреди- Потписот 7 има преовладување на C>T замени на места на соседните пиримидини (соседно на C или T), а особено дијагностично подмножесто е CC>TT динуклеотидната мутација. Овој модел се појавува затоа што големите фотопроизводи на ДНК предизвикани од ултравиолетовата светлина ги придружуваат двете соседни пиримидини; фотопроизводот обично е циклобутан пиримидин димер.[19] Специфичноста за C>T се чини дека се должи на милионскократното забрзување на деаминацијата на C кога е дел од циклобутан пиримидин димерот, со резултираниот урацил кој дејствува како T.[20][21] Циклобутан пиримидин димерите се поправани преку поправка со вадење на базата придружен со транскрипција, предизвикувајќи силна пристрасност за C>T замените збогатени на нетранскрибирана нишка на ДНК.[6] Регионите на туморско сузбивачката белковина кои се мутациски исклучени во ракот на кожа поврзан со сончевата светлина е ист како во ракот во органите кои не се изложени на сончева светлина, но мутираниот нуклеотид често е поместуван неколку бази на местото каде што може да биде направена поправка со вадење на базата.[22] Затоа, изложеноста на ултравиолетово зрачење е предложениот основен мутаген механизам на овој знак. Ултравиолетовото зрачење, исто така, ја илустрира суптилноста во толкувањето на туморскиот потпис како потпис на мутаген: само три четвртини од мутациите предизвикани од ултравиолетово зрачење во лабораторија се мутации со ултравиолетов потпис, бидејќи ултравиолетови зраци исто така предизвикува клеточни оксидативни постапки.[2] Затоа, дури и ако сите мутации во туморот биле предизвикани од ултравиолетово зрачење од сончева светлина, една четвртина од мутациите е очекувано да не се ултравиолетови мутации со потпис. Не треба да биде повикан втор канцероген за да бидат објаснети тие мутации, но потребна е втора мутациска постапка. Идентификацијата на ултравиолетов потпис во тумор со непозната првична местоположба е клинички важна бидејќи наведува дијагноза на метастазни рак на кожата и има важни импликации за третман.[23]
Алкилирачки агенси
уреди- Потписот 11 е идентификуван кај тумори кои претходно биле изложени на темозоламид, алкилирачки агенс.[6] Овој потпис е збогатен за C>T замени на гванински бази поради транскрипција поврзана со поправка со вадење на нуклеотидот. Во овој потпис е присутна силна транскрипциска пристрасност.
Тутун
уреди- И потписот 4 (пушење тутун, рак на белите дробови) и потписот 29 (џвакање тутун, гингиво-букален устен сквамозен рак) прикажуваат транскрипциона пристрасност и збогатување на C>A замените, но нивниот соодветен состав и модели (пропорција на секој вид мутација) се разликуваат малку.[6]
- Предложениот основен механизам на потписот 4 е отстранување на адукти на ДНК (тутунски бензо(а)пирен ковалентно врзан за гванин) со машината за поправка со вадење на нуклеотидот поврзана со транскрипција.[24]
Хипермутација на имуноглобулинскиот ген
уредиПотписот 9 е идентификуван кај хронична лимфоцитна леукемија и малигнен Б-клеточен лимфом и збогатување на особини за настани на трансверзија T>G. Сметано е дека е резултат на полимераза η склона кон грешки на (генот POLH) поврзан со мутагенеза.[4]
Неодамна, синтезниот потпис на полимеразата η склона кон грешка е поврзан со нехематолошки ракови (на пр. рак на кожа) и е претпоставувано дека придонесува за мутагенезата на мотивот на младиот гастричен рак и делумно може да го објасни зголемувањето на замена на динуклеотиди на TC.[25]
Историја
уредиВо текот на 1990-тите, Кертис Харис од Националниот институт за рак на Соединетите Држави, и Берт Вогелштајн од Онколошкиот центар „Џон Хопкинс“ во Балтимор ги прегледале податоците кои покажуваат дека различни видови рак имаат свој уникатен пакет на мутации во p53, кои веројатно биле предизвикани од различни агенси,[3][26] како што се хемикалиите во чадот од тутун или ултравиолетова светлина од Сонцето.[19][27] Со доаѓањето на масивното напоредно секвенционирање, Мајкл Стратон го видел потенцијалот на технологијата да го револуционизира нашето разбирање за генетските промени во поединечните тумори, поставувајќи ги огромните банки на машини за секвенционирање на ДНК на Сенгеровиот институт во движење за да ја читаат секоја буква. на ДНК во тумор.[28] До 2009 година, Стратон и неговата група ги направиле првите целосни геномски низи на рак. Тоа биле подробни карти кои ги прикажуваат сите генетски промени и мутации кои се случиле во два поединечни ракови - меланом од кожата и тумор на белите дробови.[29][30] Геномите на меланомот и ракот на белите дробови биле моќен доказ дека отпечатоците на одредени виновници може да бидат видени кај ракот со една главна причина. Овие тумори сè уште содржеле многу мутации кои не може да бидат објаснети со ултравиолетова светлина или со пушење тутун. Детективската работа станала многу посложена за рак со сложено, повеќекратно или дури и целосно непознато потекло. Како аналогија, форензичар кој брише прашина за отпечатоци од прсти на местото на убиството. Форензичарот може да има среќа и да најде сет совршени отпечатоци на прозорското стакло или рачката на вратата што одговараат на познат убиец. Сепак, многу е поголема веројатноста да откријат мешан отпечаток од прсти што им припаѓаат на цела низа луѓе - од жртва и потенцијални осомничени до невини забави и полициски истражители - сите поставени еден врз друг на секакви површини.[28] Ова е многу слично на геномите на ракот каде што повеќекратните мутациски шеми најчесто се преклопуваат еден врз друг, што ги прави податоците неразбирливи. За среќа, докторантот на Стратон, Људмил Александров смислил начин како математички да го реши проблемот. Александров покажал дека мутационите обрасци од поединечни мутагени пронајдени во туморот може да се разликуваат еден од друг со помош на математички пристап наречен раздвојување на слепи извори. Новорасположените модели на мутации биле наречени мутациски потписи.[28] Во 2013 година, Александров и Стратон ја објавиле првата сметачка рамка за дешифрирање на мутациски потписи од податоците за геномиката на ракот.[31] Потоа, тие ја примениле оваа рамка на повеќе од седум илјади геноми на ракот, создавајќи ја првата сеопфатна карта на мутациски потписи кај човечкиот рак.[32] Во моментов, повеќе од сто мутациски потписи се идентификувани низ репертоарот на човечкиот рак.[33] Во април 2022 година биле опишани 58 нови мутациски потписи.[34][35][36]
Поврзано
уредиЗабелешки
уреди- ↑ Бидејќи репликацијата, одржувањето и поправката на ДНК не е линеарна постапка, некои потписи се предизвикани од механизмите на мутагенеза кои се преклопуваат.
Наводи
уреди- ↑ 1,0 1,1 „COSMIC: somatic cancer genetics at high-resolution“. Nucleic Acids Research. 45 (D1): D777–D783. јануари 2017. doi:10.1093/nar/gkw1121. PMC 5210583. PMID 27899578.CS1-одржување: display-автори (link)
- ↑ 2,0 2,1 2,2 „UV signature mutations“. Photochemistry and Photobiology. 91 (1): 15–26. 2015. doi:10.1111/php.12377. PMC 4294947. PMID 25354245.
- ↑ 3,0 3,1 „p53 mutations in human cancers“. Science. 253 (5015): 49–53. јули 1991. Bibcode:1991Sci...253...49H. doi:10.1126/science.1905840. PMID 1905840.
- ↑ 4,00 4,01 4,02 4,03 4,04 4,05 4,06 4,07 4,08 4,09 „Signatures of mutational processes in human cancer“ (PDF). Nature. 500 (7463): 415–21. август 2013. Bibcode:2013Natur.500..415.. doi:10.1038/nature12477. PMC 3776390. PMID 23945592.CS1-одржување: display-автори (link)
- ↑ „A compendium of mutational signatures of environmental agents“. Cell. 177 (4): 821-36 e16. 2019. doi:10.1016/j.cell.2019.03.001. PMC 6506336. PMID 30982602.
- ↑ 6,0 6,1 6,2 6,3 6,4 6,5 6,6 6,7 6,8 „Clock-like mutational processes in human somatic cells“. Nature Genetics. 47 (12): 1402–7. декември 2015. doi:10.1038/ng.3441. PMC 4783858. PMID 26551669.
- ↑ „Advances in Targeted and Immunobased Therapies for Colorectal Cancer in the Genomic Era“. OncoTargets and Therapy. 9 (9): 1899–920. март 2016. doi:10.2147/OTT.S95101. PMC 4821380. PMID 27099521.
- ↑ „FDA Approval Summary: Accelerated Approval of Pembrolizumab for Second-Line Treatment of Metastatic Melanoma“. Clinical Cancer Research. 23 (19): 5666–5670. октомври 2017. doi:10.1158/1078-0432.CCR-16-0663. PMID 28235882.
- ↑ O'Neil, Nigel J.; Bailey, Melanie L.; Hieter, Philip (26 јуни 2017). „Synthetic lethality and cancer“. Nature Reviews Genetics. 18 (10): 613–623. doi:10.1038/nrg.2017.47. PMID 28649135.
- ↑ 10,0 10,1 „Homologous Recombination Deficiency and Platinum-Based Therapy Outcomes in Advanced Breast Cancer“. Clinical Cancer Research. 23 (24): 7521–7530. декември 2017. doi:10.1158/1078-0432.CCR-17-1941. PMID 29246904.CS1-одржување: display-автори (link)
- ↑ „Roles of APOBEC3A and APOBEC3B in Human Papillomavirus Infection and Disease Progression“. Viruses. 9 (8): 233. август 2017. doi:10.3390/v9080233. PMC 5580490. PMID 28825669.
- ↑ „Association of germline variants in the APOBEC3 region with cancer risk and enrichment with APOBEC-signature mutations in tumors“. Nature Genetics. 48 (11): 1330–1338. ноември 2016. doi:10.1038/ng.3670. PMC 6583788. PMID 27643540.CS1-одржување: display-автори (link)
- ↑ „Association of a germline copy number polymorphism of APOBEC3A and APOBEC3B with burden of putative APOBEC-dependent mutations in breast cancer“. Nature Genetics. 46 (5): 487–91. мај 2014. doi:10.1038/ng.2955. PMC 4137149. PMID 24728294.CS1-одржување: display-автори (link)
- ↑ „APOBEC: From mutator to editor“. Journal of Genetics and Genomics = Yi Chuan Xue Bao. 44 (9): 423–437. септември 2017. doi:10.1016/j.jgg.2017.04.009. PMID 28964683.
- ↑ „APOBEC3A and APOBEC3B Preferentially Deaminate the Lagging Strand Template during DNA Replication“. Cell Reports. 14 (6): 1273–1282. февруари 2016. doi:10.1016/j.celrep.2016.01.021. PMC 4758883. PMID 26832400.
- ↑ „A panoply of errors: polymerase proofreading domain mutations in cancer“. Nature Reviews. Cancer. 16 (2): 71–81. февруари 2016. doi:10.1038/nrc.2015.12. PMID 26822575.
- ↑ 17,0 17,1 „A Specific Mutational Signature Associated with DNA 8-Oxoguanine Persistence in MUTYH-defective Colorectal Cancer“. eBioMedicine. 20: 39–49. април 2017. doi:10.1016/j.ebiom.2017.04.022. PMC 5478212. PMID 28551381.CS1-одржување: display-автори (link)
- ↑ David, SS; O'Shea, VL; Kundu, S (2007). „Base-excision repair of oxidative DNA damage“. Nature. 447 (7147): 941–950. Bibcode:2007Natur.447..941D. doi:10.1038/nature05978. PMC 2896554. PMID 17581577.
- ↑ 19,0 19,1 „A role for sunlight in skin cancer: UV-induced p53 mutations in squamous cell carcinoma“. Proceedings of the National Academy of Sciences USA. 88 (22): 10124–8. 1991. Bibcode:1991PNAS...8810124B. doi:10.1073/pnas.88.22.10124. PMC 52880. PMID 1946433.
- ↑ „Rapid deamination of cyclobutane pyrimidine dimer photoproducts at TCG sites in a translationally and rotationally positioned nucleosome in vivo“. J Biol Chem. 290 (44): 26597–26609. 2015. doi:10.1074/jbc.M115.673301. PMC 4646317. PMID 26354431.
- ↑ „The major mechanism of melanoma mutations is based on deamination of cytosine in pyrimidine dimers as determined by circle damage sequencing“. Science Advances. 7 (31). 2021. Bibcode:2021SciA....7.6508J. doi:10.1126/sciadv.abi6508. PMC 8324051 Проверете ја вредноста
|pmc=(help). PMID 34330711 Проверете ја вредноста|pmid=(help). - ↑ „Mutation hotspots due to sunlight in the p53 gene of non-melanoma skin cancers“. Proceedings of the National Academy of Sciences USA. 90 (9): 4216–20. 1993. Bibcode:1993PNAS...90.4216Z. doi:10.1073/pnas.90.9.4216. PMC 46477. PMID 8483937.
- ↑ Mata, Douglas A.; Williams, Erik A.; Sokol, Ethan; Oxnard, Geoffrey R.; Fleischmann, Zoe; Tse, Julie Y.; Decker, Brennan (23 март 2022). „Prevalence of UV Mutational Signatures Among Cutaneous Primary Tumors“. JAMA Network Open. 5 (3): e223833. doi:10.1001/jamanetworkopen.2022.3833. PMC 8943639 Проверете ја вредноста
|pmc=(help). PMID 35319765 Проверете ја вредноста|pmid=(help). - ↑ „Mutational signatures associated with tobacco smoking in human cancer“. Science. 354 (6312): 618–622. ноември 2016. Bibcode:2016Sci...354..618A. doi:10.1126/science.aag0299. PMC 6141049. PMID 27811275.
- ↑ „DNA polymerase η mutational signatures are found in a variety of different types of cancer“. Cell Cycle. 17 (3): 348–355. февруари 2018. doi:10.1080/15384101.2017.1404208. PMC 5914734. PMID 29139326.
- ↑ „TP53 mutation spectra and load: a tool for generating hypotheses on the etiology of cancer“. IARC Scientific Publications (157): 247–70. 2004. PMID 15055300.
- ↑ „On the origin of G --> T transversions in lung cancer“. Mutation Research. 526 (1–2): 39–43. 2003. doi:10.1016/s0027-5107(03)00013-7. PMID 12714181.
- ↑ 28,0 28,1 28,2 Mosaic, Kat Arney. „The DNA detectives who are hunting the causes of cancer“. CNN. Посетено на 27 февруари 2024.
- ↑ „A comprehensive catalogue of somatic mutations from a human cancer genome“. Nature. 463 (7278): 191–6. јануари 2010. Bibcode:2010Natur.463..191P. doi:10.1038/nature08658. PMC 3145108. PMID 20016485.CS1-одржување: display-автори (link)
- ↑ „A small-cell lung cancer genome with complex signatures of tobacco exposure“. Nature. 463 (7278): 184–90. јануари 2010. Bibcode:2010Natur.463..184P. doi:10.1038/nature08629. PMC 2880489. PMID 20016488.CS1-одржување: display-автори (link)
- ↑ „Deciphering signatures of mutational processes operative in human cancer“. Cell Reports. 3 (1): 246–59. јануари 2013. doi:10.1016/j.celrep.2012.12.008. PMC 3588146. PMID 23318258.
- ↑ „Signatures of mutational processes in human cancer“. Nature. 500 (7463): 415–21. август 2013. Bibcode:2013Natur.500..415.. doi:10.1038/nature12477. PMC 3776390. PMID 23945592.CS1-одржување: display-автори (link)
- ↑ Alexandrov, Ludmil; Kim, Jaegil; Haradhvala, Nicholas J.; Huang, Mi Ni; Ng, Alvin W. T.; Boot, Arnoud; Covington, Kyle R.; Gordenin, Dmitry A.; Bergstrom, Erik (2018-05-15). „The Repertoire of Mutational Signatures in Human Cancer“. bioRxiv 10.1101/322859. Занемарен непознатиот параметар
|name-list-style=(help) - ↑ Degasperi, Andrea; и др. (21 април 2021). „Substitution mutational signatures in whole-genome–sequenced cancers in the UK population“. Science. 376 (6591). doi:10.1126/science.abl9283. PMC 7613262 Проверете ја вредноста
|pmc=(help). PMID 35949260 Проверете ја вредноста|pmid=(help). - ↑ Ledford, Heidi (2022-04-21). „Trove of tumour genomes offers clues to cancer origins“. Nature (англиски). 604 (7907): 609. Bibcode:2022Natur.604..609L. doi:10.1038/d41586-022-01095-2. PMID 35449305 Проверете ја вредноста
|pmid=(help). - ↑ „Cancer: Huge DNA analysis uncovers new clues“. BBC News (англиски). 2022-04-21. Посетено на 27 февруари 2024.