Митоза
Името митоза често се употребува како синоним за клеточната делба, меѓутоа, митозата е само делба на јадрото. Митозата, главно, се врши во телесните (соматски) клетки. Во оваа делба, јасно е видлива кончестата структура на хромозомите (од каде доаѓа и името на оваа делба - на грчки mitos значи конец). Биолошката смисла на митозата се состои во рамномерната распределба на наследниот материјал, што се содржи во хромозомите на двете новодобиени со митоза јадра. Митозата може да се смета и како и крајна видлива фаза на делење на реплицираните хромозоми.

Фази на митозата
уреди






Митозата претставува еден динамичен и континуиран краткотраен процес. Меѓутоа, за полесно објаснување на истиот, оваа делба е поделена на неколку фази, и тоа профаза, метафаза, анафаза и телофаза.
Интерфаза
уредиМитотичката фаза е релативно краток период од клеточниот циклус. Таа се одвива наизменично со многу подолгата интерфаза, во која клетката се подготвува за делба. Интерфазата е поделена на три потфази: G1 („first gap“ или „празнина“), S (синтеза) и G2 („second gap“ или „празнина“). За време на сите овие фази, клетката расте со производство на белковини и цитоплазматични органели. Меѓутоа, хромозомите се реплицираат само за време на S-фазата. На овој начин клетката расте (G1), продолжува да расте со удвојување на своите хромозоми (S), расте уште повеќе и се подготвува за митоза (G2) и на крајот се дели (M) пред повторното започнување на циклусот.[1] Сите овие потфази на интерфазата се регулирани главно преку белковини. Тие следуваат една по друга во строг редослед, а постојат и „контролни точки“ кои и даваат на клетката сигнали за премин од една во друга фаза.
Претпрофаза
уредиСамо кај растителните клетки на профазата и претходи претпрофаза. Кај растителните клетки (кои инаку поседуваат многу вакуоли) јадрото мора да мигрира во центарот на клетката пред започнувањето на митозата. Ова се постигнува со образување на фрагмозом, кој претставува напречен слој на цитоплазма кој ја пресекува клетката во пределот на рамнината каде подоцна ќе се подели клетката. Освен со образување на фрагмозомот, претпрофазата се одликува со формирање на прстен од микротубули и актински филаменти (т.н. претпрофазна лента) под плазматичната мембрана, околу екваторската рамнина на делбеното вретено кое ќе се формира подоцна. Оваа лента го означува местото каде клетката ќе се подели. Клетките на вишите растенија (на пример, цветни растенија) немаат центриоли, па затоа микротубулите образуваат вретено на самата површина на јадрото, по што се организираат во делбено вретено од самите хромозоми (откако јадрената мембрана ќе се разгради).[2] Претпрофазната лента исчезнува за време на разградувањето на јадрената обвивка и со формирањето на делбеното вретено во прометафазата.[3]
Профаза
уредиГенетичкиот материјал во јадрото се наоѓа во вид на силно расплетени нишки наречени хроматин. Со настапувањето на профазата, хроматинот се кондензира во мошне подредена структура наречена хромозом. Бидејќи генетичкиот материјал претходно се удвоил во S-фазата, реплицираните хромозоми имаат две сестрински хроматиди поврзани со центромер со помош на белковинскиот комплекс кохезин. Обично хромозомите се гледаат на големо зголемување со светлосен микроскоп.
Блиску до јадрото се наоѓаат структури наречени центрозоми кои се составени од пар на центриоли (се среќаваат само кај повеќето животински клетки). Центрозомот е координативниот центар за клеточните микротубули. Секоја клетка наследува по еден центрозом при клеточната делба, кој се реплицира со помош на јадрото пред да започне нова митоза, со што се добива пар на центрозоми. Двата центрозоми и јадрените микротубули (кои можат да се замислат како полови од кои се издаваат клеточни јажиња) го образуваат делбеното вретено со полимеризација на растворливиот тубулин. Белковините од молекуларниот мотор потоа ги придвижуваат центрозомите долж овие микротубули во насока на спротивните страни на клетката. Иако центриолите помагаат во организирање на групата од микротубули, тие не се неопходни за образување на вретеното, што може да се заклучи од тоа што отсуствуваат во растителните клетки,[2] а и од тоа што тие не секогаш се користат во митозата.[4]
Предметафаза
уредиЈадрената обвивка се разградува и микротубулите навлегуваат во јадрениот простор. Ова се нарекува отворена митоза и се среќава кај повеќето многуклеточни организми. Кај габите и некои протисти (алги или трихомонади) се забележува затворена митоза, каде вретеното се формира внатре во јадрото или неговите микротубули се способни да навлезат во интактна јадрена обвивка.[5][6]
Секој хромозом образува два кинетохори на центромерот, по еден прикачен за секоја хроматида. Кинетохорот е сложена белковинска структура што е аналогна на прстенот од куката на микротубулите; тоа е точката каде микротубулите се прикачуваат за хромозомот (околу 1-40 на број, во просек 20).[7] Иако градбата и функцијата на кинетохорите не се во целост објаснети, се знае дека во нив се наоѓа одреден вид на молекуларен мотор.[8] Кога една микротубула се поврзува со кинетохорот, моторот се активира (користејќи енергија од ATP) со што на тубулата се „искачува“ кон центрозомот од каде таа потекнува. Оваа моторичка активност, заедно со полимеризацијата и деполимеризацијата на микротубулите, ја дава придвижувачката сила потребна за одделување на двете хроматиди од хромозомот подоцна.[8]
Кога вретеното ќе ја достигне потребната должина, кинетохорните микротубули започнуваат да трагаат по кинетохори за кои ќе се закачат. Одреден број на некинетохорни микротубули заемодејствуваат со соодветните некинетохорни микротубули од спротивниот центрозом и го образуваат митотичкото вретено.[9] Прометафазата понекогаш се смета за дел од профазата. Ова е една од главните фази на митозата, бидејќи без неа не би можела да се одвива цитокинезата.
Метафаза
уредиИмето метафаза доаѓа од грчкото μετα што значи „после“. Микротубулите ги пронаоѓаат кинетохорите и се закачуваат за нив во прометафазата. Тогаш двата центрозоми почнуваат да ги привлекуваат хромозомите преку нивните закачени центромери кон двата краја на клетката. Вака хромозомите подлежат на надолжна тензија од двата краја на клетката. Може да се каже дека центромерите на хромозомите повлекуваат замислена линија долж метафазната плоча или екваторска рамнина која е на еднакво растојание од двата центрозомни полови.[9] Оваа еднаква подреденост се должи на рамнотежата на привлечните сили од спротивните кинетохори, аналогно на влечењето јаже меѓу луѓе со иста сила. Кај одредени видови на клетки хромозомите не се подредуваат на метафазна плоча, туку неодредено се движат напред-назад меѓу половите, при што само грубо се подредуваат на средната линија.
Бидејќи за правилно одделување на хромозомите треба секој кинетохор да е закачен за сноп од микротубули (нишки на вретеното), се смета дека незакачените кинетохори создаваат сигнал за да го спречат предвременото настапување на анафазата (без да се подредени сите хромозоми). Овој сигнал ја создава контролната точка на митотичкото вретено.[10]
Анафаза
уредиКога секој кинетохор е закачен за сноп од микротубули и хромозомите се подредени на метафазната плоча, клетката поминува во анафаза (од грчкото ανα што значи „горе“, „наспроти“).
Се одвиваат два настани: прво, белковините кои ги поврзуваат сестринските хроматиди сега се раскинуваат. Со тоа овие сестрински хроматиди стануваат одделени ќеркини хромозоми и се повлекуваат со скусување на кинетохорните микротубули кон соодветните центрозоми за кои претходно биле поврзани. Потоа, некинетохорните микротубули се издолжуваат, повлекувајќи ги центрозомите (како и сетот хромозоми за кои се закачени) на спротивните краеви на клетката. Силата која ги предизвикува центрозомите да се движат кон краевите на клетката е сè уште непозната, иако постои теорија која вели дека брзото создавање и разградување на микротубулите би можело да го предизвика ова движење.[11]
Овие два стадиума понекогаш се означуваат како рана и касна анафаза. Раната анафаза обично се дефинира како одделување на сестринските хроматиди, додека доцната како издолжување на микротубулите и одделување на хромозомите. На крајот од анафаза клетката ги одвоила идентичните копии на генетичкиот материјал во две одделни групации.
Телофаза
уредиТелофазата (од грч. τελος што значи „крај“) е спротивен процес на профазата и прометафазата. Во неа, некинетохорните микротубули продолжуваат да се издолжуваат, со што уште повеќе ја издолжуваат и клетката. Соодветните сестрински хромозоми се прикачуваат за спротивните краеви на клетката. Се појавува нова јадрена обвивка (од фрагментите на јадрената мембрана на клетката родител) околу секој сет на одделени сестрински хромозоми. И двата сета хромозоми, сега опкружени со нови јадра, повторно се расплетуваат во хроматин. Со ова завршува митозата, но делбата на клетката сè уште не е завршена.
Цитокинеза
уредиЦитокинезата е посебен процес кој започнува во исто време со телофазата. Кај животинските клетки се развива делбена бразда („прештипнување“) со контрактилен прстен на местото каде што била метафазната плоча, со што ги одделува одвоените јадра.[12] И кај животинските, и кај растителните клетки, делбата на клетките е исто така поттикната и од везикули добиени од Голџиевиот апарат кои се движат долж микротубулите сè до средината на клетката.[13] Кај растенијата оваа структура се претвора во клеточна плоча во центарот на фрагмопластот и се развива во клеточен ѕид, разделувајќи ги двете јадра. Фрагмопластот е микротубуларна структура карактеристична за вишите растенија, додека кај некои зелени алги се забележува фикопластен микротубуларен поредок за време на цитокинезата.[14] Секоја клетка ќерка поседува целосна копија на геномот од својата клетка родител. Крајот на цитокинезата го означува крајот на M-фазата.
Галерија
уреди-
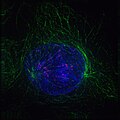 1. Профаза
1. Профаза -
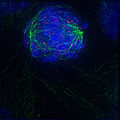 2. Прометафаза
2. Прометафаза -
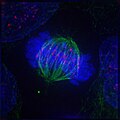 3. Метафаза
3. Метафаза -
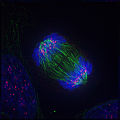 4. Анафаза
4. Анафаза -
 5. Телофаза
5. Телофаза





Наводи
уреди- ↑ Blow J, Tanaka T. (2005). „The chromosome cycle: coordinating replication and segregation: Second in the cycles review series“. EMBO Rep. 6 (11): 1028–34. doi:10.1038/sj.embor.7400557. PMC 1371039. PMID 16264427.
- ↑ 2,0 2,1 Lloyd C, Chan J. (2006). „Not so divided: the common basis of plant and animal cell division“. Nature reviews. Molecular cell biology. 7 (2): 147–52. doi:10.1038/nrm1831. PMID 16493420.
- ↑ Raven et al., 2005, pp. 58–67.
- ↑ Varmark H (2004). „Functional role of centrosomes in spindle assembly and organization“. Journal of Cellular Biochemistry. 91 (5): 904–14. doi:10.1002/jcb.20013. PMID 15034926.
- ↑ Heywood P. (1978). „Ultrastructure of mitosis in the chloromonadophycean alga Vacuolaria virescens“. Journal of Cell Science. 31: 37–51. PMID 670329.
- ↑ Ribeiro K, Pereira-Neves A, Benchimol M. (2002). „The mitotic spindle and associated membranes in the closed mitosis of trichomonads“. Biology of the Cell. 94 (3): 157–72. doi:10.1016/S0248-4900(02)01191-7. PMID 12206655.CS1-одржување: повеќе имиња: список на автори (link)
- ↑ Chan G, Liu S, Yen T. (2005). „Kinetochore structure and function“. Trends in Cell Biology. 15 (11): 589–98. doi:10.1016/j.tcb.2005.09.010. PMID 16214339.CS1-одржување: повеќе имиња: список на автори (link)
- ↑ 8,0 8,1 Maiato H, DeLuca J, Salmon E, Earnshaw W. (2004). „The dynamic kinetochore-microtubule interface“. Journal of Cell Science. 117 (Pt 23): 5461–77. doi:10.1242/jcs.01536. PMID 15509863.CS1-одржување: повеќе имиња: список на автори (link)
- ↑ 9,0 9,1 Winey M, Mamay C, O'Toole E, Mastronarde D, Giddings T, McDonald K, McIntosh J. (1995). „Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle“. Journal of Cell Biology. 129 (6): 1601–15. doi:10.1083/jcb.129.6.1601. PMC 2291174. PMID 7790357.CS1-одржување: повеќе имиња: список на автори (link)
- ↑ Chan G, Yen T. (2003). „The mitotic checkpoint: a signaling pathway that allows a single unattached kinetochore to inhibit mitotic exit“. Progress in Cell Cycle Research. 5: 431–9. PMID 14593737.
- ↑ Miller KR. (2000). „Anaphase“. Biology (5. изд.). Pearson Prentice Hall. стр. 169–70. ISBN 978-0134362656.
- ↑ Glotzer M. (2005). „The molecular requirements for cytokinesis“. Science. 307 (5716): 1735–9. doi:10.1126/science.1096896. PMID 15774750.
- ↑ Albertson R, Riggs B, Sullivan W. (2005). „Membrane traffic: a driving force in cytokinesis“. Trends in Cell Biology. 15 (2): 92–101. doi:10.1016/j.tcb.2004.12.008. PMID 15695096.CS1-одржување: повеќе имиња: список на автори (link)
- ↑ Raven et al., 2005, pp. 64–7, 328–9.
Надворешни врски
уреди| „Митоза“ на Ризницата ? |
- Анимација за митоза Архивирано на 12 јануари 2021 г. (англиски)
- Митоза (флеш анимација) (англиски)
- Видео на жива белодробна клетка на водоземец која е во фаза на митоза (англиски)
- Флеш анимација за споредба на митоза и мејоза (англиски)
- Анимација од Одделот за науки за животот при Универзитетот на Северен Тексас Архивирано на 17 март 2012 г. (англиски)
- Лекција од Академија Кан Архивирано на 28 декември 2011 г. (англиски)
- Проучување на митоза кај клетки на цицачи во култура (англиски)
- Општи К-12 училишни податоци за митоза (англиски)
- Водич за изучување на митозата за студенти Архивирано на 14 ноември 2011 г. (англиски)
- CCO Архивирано на 15 февруари 2019 г. Онтологија на клеточниот циклус (англиски)
- WormWeb.org: Интерактивна визуелизација на клеточна линија од C. elegans - Го визуелизира целото клеточно дрво и сите клеточни делби кај цевчестиот црв C. elegans (англиски)