Заднокамшични
Заднокамшични (од старогрчки ὀπίσθιος (opísthios) „задно,задни“ и κοντός (kontós) „камшик, т.е. флагела“) — широка група на еукариоти, вклучувајќи ги тука и животинското и габичното царство.[1] Заднокамшичарите, претходно наречени „група Габи/Метазои“,[2] општо се препознаени како клад. Заднокамшичарите заедно со Apusomonadida и Breviata го сочинуваат поголемиот клад Обазоа.[3][4][5][6][7]
Камшичиња и други особености
уредиВообичаена особеност на заднокамшикарите е дека камшикарните клетки, како што се сперматозоидите на повеќето животни и спорите на хитридните габи, се движат со еден заден камшик. Токму оваа карактеристика и го дава името на групата. Спротивно на тоа, камшикарните клетки во другите еукариотски групи се движат со едно или повеќе предни камшичиња. Клетките на камшичето, сепак, се последователно атрофирани во некои заднокамшикарни групи, како што е случајот со повеќето од габите.[3]
Особеностите на заднокамшикарите вклучуваат спој на вонклеточен хитин во надворешниот скелет, ѕидот на цистата/спората или клеточниот ѕид со влакновиден раст и хифи, вонклеточното варење на супстратите со осмотрофично впивање на хранливите материи и други клеточни биосинтетички и метаболички патишта. Родовите во основата на секој клад се амебоидни и фаготрофни.[8]
Историја
уредиТомас Кавалир-Смит беше првиот кој предложи дека постои поврзаност меѓу животните и габите во 1987 година,[9] и го користеше неформалното име заднокамшични (формалното име беше користено за хитридите од Коупленд во 1956 година), а беше поддржано од подоцнежните генетски истражувања.[10]
Првичните филогении ги сместувале габите во близина на растенијата и други групи кои имаат митохондрии со рамна криста, но оваа особеност е променлива. Денес се тврди дека животните и габите се многу потесно поврзани едни со други отколку што тоа би било со растенијата, бидејќи заднокамшикарите имаат тројно спојување на карбамоил фосфат синтетаза, дихидрооротаза и аспартат карбамоилтрансфераза процеси кои не се присутни во растенијата, а пак растенијата имаат спојување на тимидилат синтаза и дихидрофолат редуктаза која пак не е присутна во заднокамшикарите. Животните и габите се исто така потесно поврзани со амебите за разлика од растенијата, а растенијата се потесно поврзани со SAR супергрупата на протисти отколку со животните и габите.Животните и габите се истовремено и хетеротрофи, за разлика од растенијата, иако габите се неподвижни како растенијата, постојат и неподвижни животни.
Кавалир Смит и Стехман тврдат дека едноцилијарните еукариоти, како што се заднокамшикарите и амебозоите, заедно познати како униконти, и се одвоиле од другите двоцилијарни еукариоти, наречени биконти, набргу откако еволуирале.[11]
Таксономија
уредиЗаднокамшикарите се поделени на холомикоти (габи и сите организми поблиску поврзани со габите отколку со животните) и целовити (животни и сите организми поблиску поврзани со животните отколку со габите), сè уште не е препознаен заднокамшикар заснован на поделбата холимикота/целовит. Заднокамшикарите во голема мера беа распознаени и поделени од страна на Ториела и сор.[12] Холимикота и целовитите организми се составени од следните групи.
- Холомикота (налик на габа)
- Габи
- Вклучува:
- хитриди (флагелирани, зооспорични габи)
- Fonticula [13] ( денес се разгледуваат како дел од Cristidiscoidea, сестринска група на габите)
- Хијалорафидиум (претходно се разгледувала како зелена алга, сега се смета за габа)
- Микроспоридија (претходно се разгледувала како апикомплексија )
- Nucleariida ( денес се разгледуваат како дел од Cristidiscoidea, сестринска група на габите)
- Исклучува:
- лавиринтуломицети (лигавомрежни) (сега вклучени во супергрупата SAR )
- миксомицети (сега вклучени во амебозои )
- оомицети (водени калапи) (сега вклучени во супергрупата SAR )
- Вклучува:
- Розелида (не е утврдена припадноста)
- Габи
- Целовити (налик на животно)
- Коралохитриум (порано се разгледувал како хетероконт)
- Кончести
- Животни (вклучувајќи миксозои )
- Околувратни камшикари (камшикари порано разгледувани како протозои)
- Ѕвездокончести
- Мезомицетозои
- Амебидијали (порано се разгледуваа како трихомицети)
- Dermocystida (порано се разгледуваа како паразитски габи или спорозоа)
- Eccrinales (порано се разгледуваа како габи)
- Ichthyophonida (порано се разгледуваа како паразитски габи incertae sedis )
Филогенија
уредиСледното филогенетско стебло укажува на еволутивните односи помеѓу различните опистоконтски лоза и временската дивергенција на кладите пред милиони години (Mya). [14][15][16]
| Amorphea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Галерија
уреди-

-

-
 Микроспоридијална спора (Микроспоридија)
Микроспоридијална спора (Микроспоридија) -
 Хитрид(флагелирана габа)
Хитрид(флагелирана габа) -
 Сфаероформасп. (Мезомицетозои)
Сфаероформасп. (Мезомицетозои) -
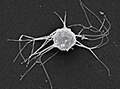
-
 Капсаспорасп. (Ѕвездокончести)
Капсаспорасп. (Ѕвездокончести) -

-
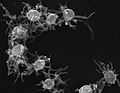
 Дезмареласп. колонија (Околувратни камшикари)
Дезмареласп. колонија (Околувратни камшикари) -











Наводи
уреди- ↑ „Multigene phylogeny of choanozoa and the origin of animals“. PLOS ONE. 3 (5): e2098. May 2008. Bibcode:2008PLoSO...3.2098S. doi:10.1371/journal.pone.0002098. PMC 2346548. PMID 18461162.
- ↑ „Fungi/Metazoa group“. UniProt. Архивирано од изворникот на 17 February 2009. Посетено на 2009-03-08.
- ↑ 3,0 3,1 „The protistan origins of animals and fungi“. Molecular Biology and Evolution. 23 (1): 93–106. January 2006. doi:10.1093/molbev/msj011. PMID 16151185.
- ↑ Huang, Jinling; Xu, Ying; Gogarten, Johann Peter (November 2005). „The presence of a haloarchaeal type tyrosyl-tRNA synthetase marks the opisthokonts as monophyletic“. Molecular Biology and Evolution. 22 (11): 2142–2146. doi:10.1093/molbev/msi221. PMID 16049196.
- ↑ „Evaluating support for the current classification of eukaryotic diversity“. PLOS Genetics. 2 (12): e220. December 2006. doi:10.1371/journal.pgen.0020220. PMC 1713255. PMID 17194223.
- ↑ Torruella, G.; Derelle, R.; Paps, J.; Lang, B. F.; Roger, A. J.; Shalchian-Tabrizi, K.; Ruiz-Trillo, I. (February 2012). „Phylogenetic relationships within the Opisthokonta based on phylogenomic analyses of conserved single-copy protein domains“. Molecular Biology and Evolution. 29 (2): 531–544. doi:10.1093/molbev/msr185. PMC 3350318. PMID 21771718.
- ↑ Eme, L.; Sharpe, S. C.; Brown, M. W.; Roger, A. J. (August 2014). „On the age of eukaryotes: evaluating evidence from fossils and molecular clocks“. Cold Spring Harbor Perspectives in Biology. 6 (8): a016139. doi:10.1101/cshperspect.a016139. PMC 4107988. PMID 25085908.
- ↑ Adl, Sina M.; Bass, David; Lane, Christopher E.; Lukeš, Julius; Schoch, Conrad L.; Smirnov, Alexey; и др. (January 2019). „Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes“. The Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi:10.1111/jeu.12691. PMC 6492006. PMID 30257078.
- ↑ Rayner, Alan D. M., уред. (1987). „The origin of fungi and pseudofungi“. Evolutionary biology of Fungi. Cambridge: Cambridge University Press. стр. 339–353. ISBN 0-521-33050-5.
- ↑ „Monophyletic origins of the metazoa: an evolutionary link with fungi“. Science. 260 (5106): 340–342. April 1993. Bibcode:1993Sci...260..340W. doi:10.1126/science.8469985. PMID 8469985.
- ↑ Stechmann, Alexandra; Cavalier-Smith, Thomas (July 2002). „Rooting the eukaryote tree by using a derived gene fusion“. Science. 297 (5578): 89–91. Bibcode:2002Sci...297...89S. doi:10.1126/science.1071196. PMID 12098695.
- ↑ Torruella, Guifré; De Mendoza, Alex; Grau-Bové, Xavier; Antó, Meritxell; Chaplin, Mark A.; Del Campo, Javier; и др. (September 2015). „Phylogenomics Reveals Convergent Evolution of Lifestyles in Close Relatives of Animals and Fungi“. Current Biology. 25 (18): 2404–2410. doi:10.1016/j.cub.2015.07.053. PMID 26365255.
- ↑ Brown, M. W.; Spiegel, F. W.; Silberman, J. D. (December 2009). „Phylogeny of the "forgotten" cellular slime mold, Fonticula alba, reveals a key evolutionary branch within Opisthokonta“. Molecular Biology and Evolution. 26 (12): 2699–2709. doi:10.1093/molbev/msp185. PMID 19692665.
- ↑ Tedersoo, Leho; Sánchez-Ramírez, Santiago; Kõljalg, Urmas; Bahram, Mohammad; Döring, Markus; Schigel, Dmitry; и др. (2018). „High-level classification of the Fungi and a tool for evolutionary ecological analyses“. Fungal Diversity. 90 (1): 135–159. doi:10.1007/s13225-018-0401-0. ISSN 1560-2745.
- ↑ Galindo, Luis Javier; Torruella, Guifré; López-García, Purificación; Ciobanu, Maria; Gutiérrez-Preciado, Ana; Karpov, Sergey A.; Moreira, David (June 2023). „Phylogenomics Supports the Monophyly of Aphelids and Fungi and Identifies New Molecular Synapomorphies“. Systematic Biology. 72 (3): 505–515. doi:10.1093/sysbio/syac054. PMID 35900180 Проверете ја вредноста
|pmid=(help). - ↑ Tikhonenkov, Denis V.; Mikhailov, Kirill V.; Hehenberger, Elisabeth; Karpov, Sergei A.; Prokina, Kristina I.; Esaulov, Anton S.; и др. (November 2020). „New Lineage of Microbial Predators Adds Complexity to Reconstructing the Evolutionary Origin of Animals“. Current Biology. 30 (22): 4500–4509.e5. doi:10.1016/j.cub.2020.08.061. PMID 32976804.